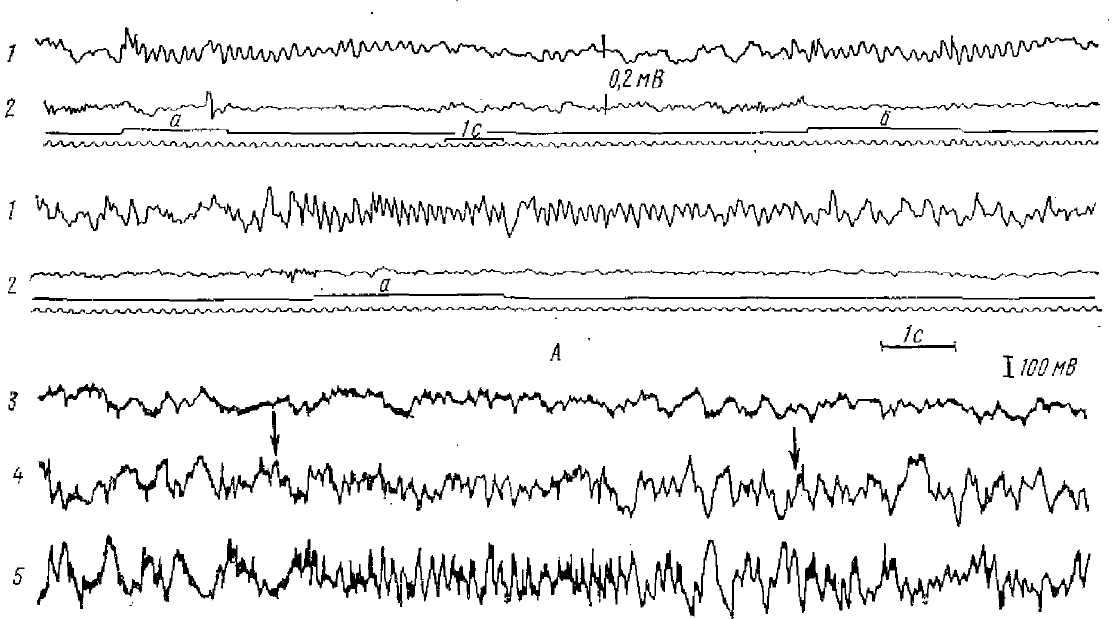
A — первые записи θ -активности (1) гиппокампа у кролика при болевом раздражении лапы (а) и хвоста (б); 2 — контрольная запись с хвостатого ядра (но Юнгу и Корнмюллеру, 1938). Б — θ-ритм, вызванный у морской свинки на присутствие экспериментатора в камере (отмечено стрелками), 3 — кош рольная запись с неокортскса; 4, 5 — активность гиппокампа (по Либерсону и Кадилаку 1953)
➥ Основная статья: Тета-ритмы
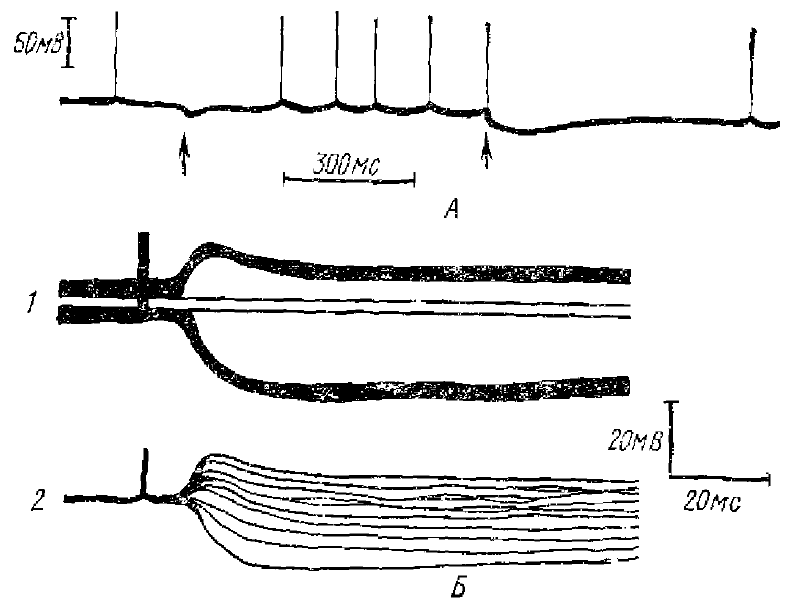
Многочисленными исследованиями установлено, что θ-ритм возникает в электрограмме ряда структур мозга млекопитающих при безусловнорефлекторной и условнорефлекторной деятельности и является одним из компонентов ориентировочной реакции животного (Green, Arduini, 1954; Эйди, 1963). Наиболее действенны при вызове θ-ритма болевые, проприоцептивные раздражители, менее — обонятельные, слуховые и световые (Petsche, Stumpf, 1962; Дыкман, 1964 и многие другие) (рис. 1).
Другой термин θ-ритма — стресс-ритм, или ритм напряжения, — свидетельствует о том, что этот ритм проявляется в биологически неблагоприятных ситуациях, при состоянии напряжения, которое вызывается агентами разной природы. Это послужило основанием считать его биоэлектрическим выражением биологически отрицательной реакции (Анохин, 1962). Хотя эта точка зрения подкреплена обширным экспериментальным материалом, разделяют ее не все авторы. Изучение ЭЭГ кролика в условиях свободного поведения (Чжоу Шао-цы, 1958; Шумилина, 1961) показало, что в этом случае почти вся фоновая запись представляет собой непрерывную θ-активность, в то время как θ-ритм в ЭЭГ фиксированного животного возникает, как правило, лишь в ответ на раздражения (Новикова, Фарбер, 1959, и др.). Иными словами, фиксация животного, являющаяся для него биологически неблагоприятной ситуацией, слабее активирует θ-ритм, чем пребывание животного в экспериментальной камере в нефиксированном состоянии. Результаты исследований многих авторов на кроликах (Шумилина, 1961; Воронин, Котляр, 1962; Эйди, 1963; Захарова, Калюжный, 1965, и др.) показывают, что появление θ-ритма сопутствует и положительным пищевым реакциям животного. Однако у крыс и кошек он коррелирует, как правило, с биологически отрицательной ситуацией.
Ряд исследователей полагают, что θ-ритм отражает правильное решение задачи животным (Elazar, 1967; Holmes et al., 1969), активацию нервных механизмов произвольных движений (Paxinos et ah, 1969, 1970), ЭЭГ проявление пищевых, питьевых и оборонительных эмоциональных реакций (Ониаyи и др., 1972), ориентировочно-исследовательскую реакцию, связанную с движением вибрисс (опыты на крысах) (Gray, 1970) и т. д. Эйди (Adey et al., 1960) выдвинул гипотезу о том, что появление θ-ритма во время обучения отражает роль гиппокампа в процессах принятия решения и консолидации памяти. Однако эта гипотеза была подвергнута сомнению (Bennet, Gotteried, 1969).
Большинство авторов полагают, что во время обучения θ-ритм гиппокампа является коррелятом ориентировочной реакции, возникающей на отмену привычного подкрепления (Gray, 1971). Граштпан и сотрудники (Gras- tyan et al., 1966) считают, что θ-ритм сопровождает лишь ориентировочную реакцию животного, а условнорефлекторная деятельность (при прочной условной связи) вызывает только его десинхронизацию; появление θ-ритма во время условного рефлекса свидетельствует о растормаживании ориентировочной реакции.
Наибольшую амплитуду θ-ритм имеет в гиппокампе, он отмечен также в неспецифических структурах среднего и промежуточного мозга, в своде, в маммилярных телах. В энторинальной, затылочной и теменно-височной коре кролика реакция пробуждения выражается в виде θ-ритма, тогда как в лобной и сенсомоторной коре он регистрируется реже, и реакция пробуждения выражается обычно в виде реакции десинхронизации. Исследование θ-ритма неанестезированного кролика топоскопическим методом (Petsche, Sttimpf, 1960) дало возможность сравнить соотношение фаз θ-активности в нескольких точках различных отделов мозга. Обработка результатов показала, что фронт волны θ-ритма распространяется в виде сферической волны из точки, совпадающей с анатомическим расположением перегородки (septum). Скорость распространения оказалась равной 20—80 см в 1 с. Объяснения такому распространению не найдены (см. Гусельников, Супин, 1968).
Исходя из того, что θ-ритм в новой коре оказывается наложенным на быстрые низковольтные колебания и наибольшая регулярность и наивысшая амплитуда его наблюдается в задних отделах новой коры, т. е. в областях, расположенных над гиппокампом, Петче и Штумпф заключили, что θ-ритм в новой коре, это пассивное электрическое явление, — следствие распространения электрического поля потенциалов гиппокампа. Грин с соавторами (Green et al,, 1960) показали, что θ-ритм новой коры находится в постоянном фазовом соотношении с θ-ритмом гиппокампа. Они сдвинуты по фазе на 180°, если регистрируются потенциалы точки гиппокампа, лежащей в области апикальных дендритов пирамидных клеток, и синфазны при положении гиппокампального электрода в области их базальных дендритов. Таким образом, по мнению авторов, новая кора не является активным генератором θ-ритма, а колебания корковой биоэлектрической активности в ритме гиппокампального θ-ритма представляют своего рода артефакт.
Однако в ряде случаев, как в остром, так и в хроническом эксперименте, генерация θ-ритма новой коры может быть обнаружена. Так, найдено, что повреждение centrum medianum блокирует его в дорзальном гиппокампе при сохранении вспышек в новой коре, а разрушение дорзомедиального ядра таламуса выключает его в поясной извилине с сохранением в гиппокампе.
После коагуляции ряда неспецифических ядер таламуса θ-ритм исчезал в новой коре, но сохранялся в гиппокампе. Показано также, что амплитуда волн его в попой коре кошки уменьшается при локальном охлаждении или кокаинизации обнаженной коры. Функциональное выключение гиппокампа крыс методом распространяющейся депрессии (Weiss, 1961) не устраняет θ-ритм в ЭЭГ новой коры. Очевидно, если бы корковый θ-ритм всецело определялся гиппокампальными колебаниями потенциала, то все те воздействия, из-за которых он исчезал в гиппокампе, должны были бы непременно устранять его и из корковых отведений, чего в действительности не наблюдается.
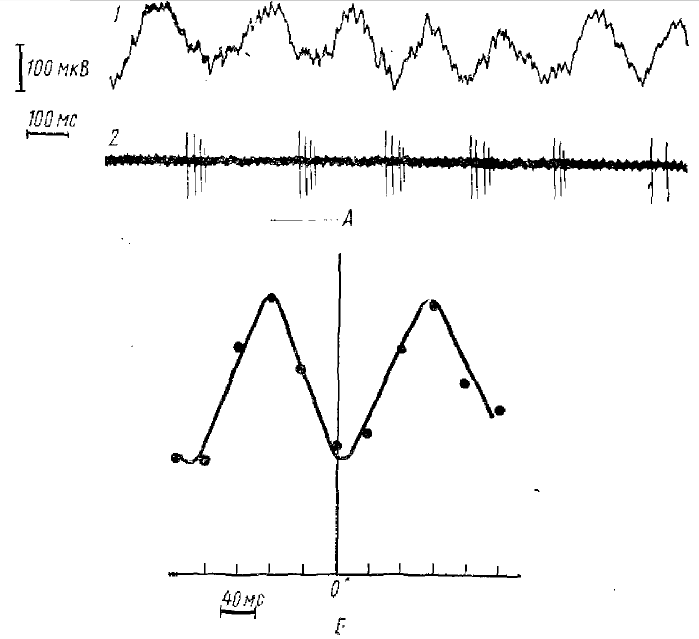
По ординате — отклонение линии медленной активности от условной нулевой линии в относительных единицах. Положительная полярность — вверх. Использован корреляционный метод. Для каждого нейрона бралось 50 импульсных разрядов подряд во время θ -ритма. Анализу подвергались участки электрограмм (ЭГ) с θ -ритмом до 490 мс (200 мс до разряда и 200 мс после разряда клетки). Анализ состоит в следующем; с интервалом в 20 мс намерялись отклонения точек ЭГ θ-ритма от условной нулевой линии, затем высчитывались средние арифметические для соответствующих точек 50 участков θ-ритма. По 21 Полученной средней величине строилась кривая — коррелограмма импульсных разрядов нейрона и θ-ритма. Считалось, что между этими процессами существует связь, если форма коррелограммы напоминала волну θ-ритка и разница между ее максимумами и минимумами была достоверна с вероятностью 0,95
Исследования (Супин, 1964), посвященные анализу взаимодействия вызванных потенциалов с разными формами фоновой корковой ЭЭГ кролика, показали, что амплитуда положительной фазы первичного ответа на вспышку света оказывается несколько больше в том случае, если она приходится на отрицательную фазу θ-волны (разница статистически достоверна). Г. В. Георгиевым (1966) было найдено, что около половины (28 из 50) нейронов неокортекса неанестезированного кролика обнаруживают достоверную корреляцию с. определенными фазами θ-ритма (рис. 2). Взаимодействие волн θ-ритма с такими корковыми формами электрической активности, как первичный ответ зрительной коры и разряды корковых нейронов, не может не свидетельствовать о генерации его волн элементами коры.
Изучая корреляционным методом биоэлектрическую активность мозга хронически оперированных кошек, Эйди с сотрудниками (Adey at al., 1960) показали, что в процессе выработки пищедобывательных движений в простом лабиринте происходит изменение соотношения фаз θ-ритма гиппокампа и энторинальной коры. Подобная подвижность фазовых соотношений θ-ритма в таких разных нервных структурах может свидетельствовать о различных анатомических путях, вызывающих его, На активную роль коры в генерации коркового θ-ритма указывают и данные других исследователей.
Т. А. Корольковой и В. Д. Трут (1971) на необездвиженных кроликах в хронических условиях опыта с использованием специальных методов анализа результатов было выяснено, что в – спокойном состоянии животного и без применения каких-либо радражений θ-ритм в новой коре есть чисто физическое отражение электрических полей, генерируемых гиппокампом. Но при специальных воздействиях, вызывающих функционально значимое повышение синхронности корковых потенциалов θ-ритма, было выяснено, что в этом случае такая синхронность определяется чисто корковыми процессами. Однако отсутствие окончательной ясности в этой проблеме требует особой осторожности в функциональной оценке корковой синхронизированной активности пробуждения и особенно в трактовке экспериментов с ориентировочными и условными реакциями животного.
Как упоминалось выше, телескопическое исследование Петче и Штумпфа распространения θ-ритма кролика показало, что исходная точка сферической волны его топографически совпадает с перегородкой (septum). Еще ранее Грин и Ардуини (Green, Arduini, 1954) в хронических опытах на кроликах, продолжавшихся до 30 дней, отметили, что электролитическое разрушение перегородки необратимо устраняет θ-ритм из всех отведений, причем реакция пробуждения в гиппокампе принимает характер десинхронизации. Брюкке и Майер с сотрудниками (Briicke et al., 1959; Mayor, Stumpt, 1958) и в нашей лаборатории И. И. Полетаева (1968), подтвердив этот факт, показали, что локальная инъекция малых доз прокаина в область перегородки подавляет θ-ритм, но в дальнейшем он полностью восстанавливается. Исчезновение его после разрушения перегородки наблюдали также у крыс.
Последующими работами установлено, что роль перегородки сводится к роли водителя ритма (Вгuсke, 1959). Залпы активности одиночных нейронов перегородки четко коррелируют с волнами θ-ритма гиппокампа (Petsche et al., 1962). Увеличение частоты его при усилении раздражающего влияния (фармакологического воздействия или раздражения среднемозговой ретикулярной формации) происходит всегда параллельно с учащением залпов разрядов нейронов перегородки и увеличением числа спайков в разряде.
Дальнейшее исследование, септального водителя θ-ритма показало, что средний латентный период начала нейронного залпа неодинаков для разных нейронов, однако клетки, расположенные в непосредственной близости друг от друга, обычно разряжаются со сходными латентными периодами. Кроме того, внутри этой структуры существуют некоторые немногочисленные популяции клеток. Нейроны каждой такой группы разряжаются синхронно друг с другом и асинхронно с нейронами иных совокупностей. Одна из задач водителя ритма заключается в превращении непрерывного потока импульсов, поднимающегося из ретикулярной формации среднего мозга, в прерывистые залпы разрядов, которые затем передаются в гиппокамп.
Опытами Гоголака и др. (Gogolak et al., 1967) было выяснено, что не все нейроны перегородки, а только нейроны дорзального отдела (диагональной полоски Брока) работают как пейсмейкер гиппокампального θ-ритма. Большинство из них разряжаются пачками, синхронными с θ-волнами. Результаты опытов также показали, что θ-ритм возникает только в том случае, если около 80% нейронов этого участка разряжаются пачками.
Авторы отмечают, что основным фактором в провокации θ-ритма является общее число спайковых разрядов, получаемых гиппокампом, т. е. уровень синаптической бомбардировки в каждый данный момент, а не фазовые соотношения каждой пачки с θ-волнами.
Причина строгой ритмичности разрядов клеток перегородки кроется, по-видимому, в их функциональных особенностях. Во всяком случае, анализ длительности залпа и длительности периода θ-ритма показал, что между этими величинами существует строгая линейная зависимость: при увеличении частоты θ-ритма (уменьшения периода θ-волны) уменьшается длительность залпа. Авторы предположили, что тормозные влияния, ограничивающие разряд нейронов, создаются, видимо, коллатеральным торможением. Какова бы ни была тонкая организация ритмической импульсации перегородки, возбуждающее влияние, которое она оказывает на гиппокампальные структуры, является необходимым условием генерации θ-ритма в гиппокампе.
В экспериментах Брюкке и др. (Briicke et al., 1959) было найдено, что низкочастотное раздражение перегородки вызывает в ряде структур мозга ритмическую реакцию, которая обнаруживает сходство со спонтанным θ-ритмом по многим признакам; форме, амплитуде отдельных волн, а также распространению по структурам мозга. В пользу сходства наблюдаемой ритмической реакции со спонтанным θ-ритмом говорит и то, что реакция возникает при раздражении перегородки, т. е. первичного очага стресс-ритма.
Волны типа θ-ритм могут следовать за ритмическими раздражениями перегородки в довольно широком диапазоне частот. Раздражение перегородки с высокой частотой (100 имп/с) ритмической реакции не вызывает; более того, спонтанный θ-ритм блокируется во всех структурах и замещается нерегулярной высокочастотной активностью. Наиболее высокая амплитуда волн ритмической реакции возникает при частоте раздражений, близкой к частоте спонтанного ритма. Анализ показал, что эта реакция возникает путем запуска каждым стимулом новой медленной волны на фоне предыдущей волны, причем при некоторых частотах раздражения наблюдается временная суммация волн. Таким образом, θ-ритм может быть искусственно вызвав или видоизменен путем ритмических раздражений первичного очага этой активности. Преобразование ритмических сигналов перегородки в θ-волны гиппокампа определяется, по-видимому, особенностями нейронной организации гиппокампа.
Что касается путей следования афферентов перегородки к пирамидным нейронам гиппокампа, то они еще изучены недостаточно, Считается, что афференты перегородки оканчиваются на клетках зубчатой фасции, причем моховидные волокна последних образуют контакты с пирамидными клетками 3 и 4 полей гиппокампа, а шафферовские коллатерали этих нейронов вступают в контакты с пирамидными клетками 1 и 2 полей.
Однако существует также мнение, что афференты перегородки могут прямо оканчиваться на апикальных дендритах пирамид 3 и 4 полей, т. е. путь от перегородки к пирамидам 1 и 2 полей может иметь одно синаптическое переключение.
Ритмическая активность нейронов гиппокампа
Уже в работах Грина и Макис (Green, Machne, 1955) было отмечено, что спонтанная импульсная активность нейронов гиппокампа может иметь вид сгруппированных пачек импульсов, следующих в такте с суммарной ритмической электрической активностью.
Более детальные исследования (Green et al., 1961) показали, что импульсные разряды пирамидных клеток гиппокампа и суммарный θ-ритм, регистрируемые с одного и того же микроэлектрода, в большинстве случаев связаны между собой: импульсы совпадают с фазой перехода от негативной к позитивной вершине волны; этим свойством обладает около 3/4 всех пирамидных нейронов. Очевидно, это указывает на тесную связь процессов в отдельных нейронах гиппокампа с его суммарной электрической активностью — θ-ритмом. Детальные исследования внутриклеточных колебаний потенциала в нейронах гиппокампа и экстраклеточного распределения токов в гиппокампе во время θ-ритма позволили установить, какие именно процессы в нейронах связаны с θ-ритмом и ответственны за его генерацию.
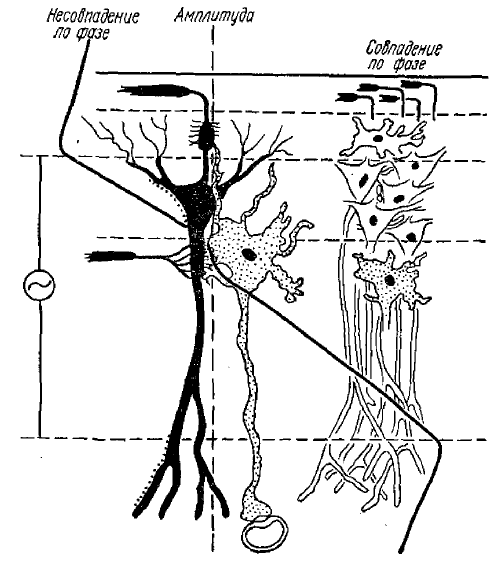
Жирная линия — амплитуда нанесенной на рисунок, θ -волны проходит через нулевой потенциал, т. е. меняется на обратную в области проксимальной части апикального дендрита
Исследование распределения потенциалов θ-ритма по слоям гиппокампа выявило существование четкой дипольной картины (рис. 3). θ-ритм имеет максимальную амплитуду (но противоположную полярность) в слое между телами и базальными дендритами пирамидных нейронов и в слое между телами и разветвлениями апикальных дендритов этих нейронов (Green et al., 1959, 1961; Green, 1962). Между этими слоями наблюдается картина дипольного переворота потенциалов. При передвижении регистрирующего электрода вдоль оси пирамидных нейронов гиппокампа амплитуда волн θ-ритма уменьшается без изменения фазы (т. е. волны остаются синхронными с волнами, регистрируемыми от поверхности гиппокампа), затем амплитуда волн доходит практически до нуля (несколько глубже слоя тел пирамидных нейронов), а затем появляется и нарастает по амплитуде активность, являющаяся практически зеркальным отражением активности, регистрируемой по другую сторону уровня инверсии (нулевой амплитуды). Эти данные позволили заключить, что θ-ритм гиппокампа генерируется не распространяющимися процессами типа потенциалов действия, а локальными распространяющимися процессами, скорее всего постсинаптическими потенциалами.
Такое представление о природе θ-ритма гиппокампа Подтверждается и данными внутриклеточной регистрации потенциалов. Детальное исследование внутриклеточной активности пирамидных нейронов гиппокампа в сопоставлении с θ-ритмом показало, что у большинства нейронов можно наблюдать ритмические колебания внутриклеточного потенциала , имеющие значительную амплитуду (около 12 мВ).
➥ Более подробно: Гипотезы о природе суммарной электрической активности головного мозга
Они вызывают соответствующие ритмические изменения интенсивности импульсной активности нейронов: на деполяризационной фазе колебаний внутриклеточного потенциала наблюдаются импульсные разряды, на гиперполяризационной фазе разряды отсутствуют. Между θ-ритмом гиппокампа и внутриклеточными колебаниями потенциала наблюдается полное соответствие не только по форме и временным характеристикам, но и по их изменениям при афферентных раздражениях (при нарушении связей перегородки с гиппокампом исчезает и суммарный и внутриклеточный θ-ритм; Шабан, 1969, 1970). Из этого следует, что суммарная электрическая активность гиппокампа и соответствующие ей колебания внутриклеточного потенциала пирамидных нейронов непосредственно связаны между собой. Иными словами, экстраклеточный ритмический θ-ритм гиппокампа и ритмические колебания внутриклеточного потенциала пирамидных нейронов, по-видимомy, являются следствием одного и того же процесса — синаптических токов пирамидных нейронов.
Какие же группы синапсов генерируют токи, вызывающие ритмические колебания внутриклеточных потенциалов пирамидных нейронов и экстраклеточных потенциалов в гиппокампе? Изложенные результаты распределения экстраклеточных потенциалов показывают, что один из полюсов генератора θ-ритма расположен между телами и базальными дендритами пирамидных нейронов гиппокампа, а другой — в области апикальных дендритов. Очевидно, расположение синапсов, генерирующих θ-ритм, должно совпадать с положением одного из этих полюсов; другой может возникать вследствие пассивного входа в нейрон или выхода из нейрона тока, генерируемого в области расположения синапсов.
Как показывают исследования вызванных потенциалов гиппокампа на раздражение перегородки (Andersen et, al., 1961) и соотношения вызванных потенциалов с импульсной активностью нейронов (Briicke et al., 1963), первичным эффектом в пирамидных нейронах гиппокампа при приходе импульсов из перегородки должны быть ВПСП в области базальных частей апикальных дендритов. Этот ВПСП сопровождается первоначальным повышением импульсной активности пирамидных нейронов гиппокампа.

Пирамидный нейрон возбуждается одиночным раздражением. Внутриклеточная регистрации. Момент раздражения отмечен артефактом
Однако изменения внутриклеточного потенциала пирамидных нейронов не ограничиваются первичным ВПСП. Характерной чертой реакции пирамидных нейронов гиппокампа является довольно длительное (десятки миллисекунд) высокоамплитудное деполяризационное колебание потенциала (Euler, Green, 1960; Kandel, Spencer, 1961). Эта глубокая деполяризация складывается, очевидно, из следовых деполяризационных колебаний относительно небольшой амплитуды (около 10 мВ), наблюдаемых после одиночных импульсных разрядов пирамидных нейронов гиппокампа и обнаруживающих способность к суммации при появлении нескольких импульсных разрядов (Kandel, Spencer, 1961). Следовая деполяризация после каждого импульса облегчает появление следующего импульса; сопровождающая его следовая деполяризация суммируется с первой волной деполяризации, еще больше облегчая возможность появления следующих импульсов, и т. д. Возникает своего рода положительная обратная связь, приводящая к быстрому росту деполяризации и к появлению высокочастотных импульсных разрядов на ее фоне. Однако нарастание этого процесса происходит недолго, глубокая деполяризация нейронов приводит к угнетению генерации импульсной активности по типу католической депрессии (рис. 4).
По мере нарастания деполяризации амплитуда импульсных разрядов нейрона уменьшается, и вскоре они совсем прекращаются (рис. 4, Д). Таким образом, процесс суммирования и нарастания деполяризации приводит к появлению высокочастотной, но относительно короткой (обычно всего из нескольких разрядов) вспышки импульсной активности пирамидного нейрона. Этот процесс получил название «процесса инактивации». После прекращения импульсных разрядов пирамидных Нейронов деполяризация в них перестает поддерживаться и спадает. Формирующиеся таким образом деполяризационные колебания отражаются в позитивных колебаниях потенциала, регистрируемого от дорзальной поверхности гиппокампа; импульсные разряды пирамидных нейронов соответствуют восходящей фазе (переходу к позитивной вершине) этих суммарных экстраклеточных колебаний.
Поскольку волны глубокой деполяризации (процессы инактивации) являются характерной чертой реакции пирамидных нейронов гиппокампа на раздражения (в том числе на раздражение перегородки), естественно ожидать, что они будут возникать и под влиянием залпов импульсов из перегородки во время θ-ритма, играя существенную роль в формировании регистрируемых от гиппокампа потенциалов θ-ритма. В пользу этого говорит одинаковое соотношение θ-ритма и экстраклеточных потенциалов, сопровождающих процесс инактивации, с импульсной активностью пирамидных нейронов.
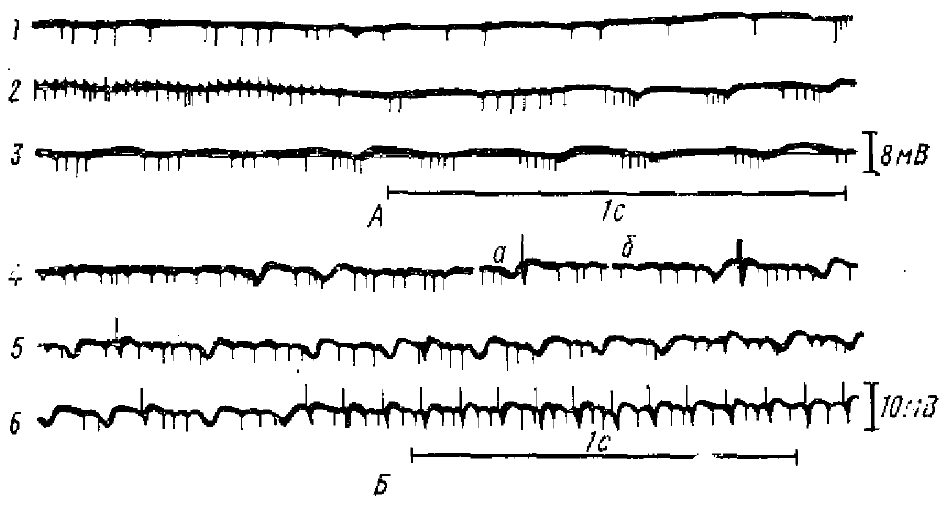
А — экстраклеточная импульсная я медленная активность до стимуляции (1), во время (2) и после повторных раздражений перегородки (3): 2 и 3—последовательные отрезки непрерывной записи (инактивационные процессы в экстраклеточном отведении выражаются в виде позитивных волн, сопровождающихся уменьшением амплитуды разрядов нейрона); Б — эффект раздражения перегородки короткими сериями импульсов: 4— контроль, а и б — записи во время раздражений, 5, 6 — ритмическая серия процессов инактивации, возникающая после раздражений. Одиночные раздражения перегородки (отмечены из 4 и 5 артефактами) нарушают последовательность волн инактивации
Особенно заметно сходство θ-ритма с процессами инактивации, если последние вызываются ритмическим раздражением перегородки. Возникающая при этом серит волн инактивации с вспышками импульсной активности на фазе нарастания каждой позитивной волны (рис. 5) чрезвычайно сходна со спонтанным θ-ритмом (Euler, 1960; Euler, Green, 1960).
Пока по вполне ясно, какова природа деполяризационных колебаний, из которых складывается волна глубокой деполяризации. Наиболее вероятное представление, что деполяризационные колебания являются ВПСП. Эти ВПСП могут возникать в результате разрядов самих же пирамидных нейронов гиппокампа через посредство системы возвратных возбуждающих связей, например, коллатералей аксонов, пирамидных нейронов. Возможность такого процесса подтверждается тем, что деполяризационные колебания типа ВПСП могут возникать в пирамидных нейронах гиппокампа и при антидромном их возбуждении (Spencer, Kandеl, 1962). Такая система возвратного возбуждения (положительная обратная связь) вполне может обеспечить лавинообразное нарастание деполяризации пирамидных нейронов и их импульсной активности, которая наблюдается на восходящей фазе волны инактивации.
1 — нормальный ТПСП (нижняя кривая) и результат инверсии ТПСП (верхняя кривая); 2 — последовательность стадий инверсии ТПСП
Процессы инактивации — не единственный вид относительно медленных процессов, которые могут принимать участие в генерации суммарного θ-ритма. Как показывают данные Кэндела и др. (Kandel et al., 1961), Спенсера и Кэндела (Spencer, Kandel, 1962) разряды пирамидных нейронов гиппокампа могут сопровождаться последующим дополнительным гиперполяризационным потенциалом значительной амплитуды (рис. 6). Этот гиперполяризационный потенциал может возникать при антидромной активации пирамидных нейронов после антидромного спайка и следовой деполяризации. Появление антидромного спайка не обязательно для возникновения длительной гиперполяризации при слабых антидромных раздражениях антидромный разряд может не наблюдаться, но гиперполяризация возникает. Эти факты, а также инверсия ТПСП при диффузии в нейрон КСl (рис. 6, Б, 1, 2) позволяют сделать заключение о природе наблюдаемой гиперполяризации: она является ТПСП, возникающим в пирамидных нейронах гиппокампа через посредство возвратных коллатералей их аксонов. Детальные исследования Андерсена и др. (Andersen et al., 1963, 1964) подтвердили существование в гиппокампе системы возвратного коллатерального торможения и позволили уточнить структуру этой системы (см. лекцию 2). Активация тормозных синапсов на пирамидных нейронах сопровождается гиперполяризацией нейронов и длительным позитивным колебанием экстраклеточного потенциала на уровне тел пирамидных нейронов. В области апикальных дендритов пирамидных нейронов при этом регистрируется отраженная негативность.
Если учесть значительную амплитуду возвратных ТПСП и соответствующих им позитивных колебаний экстраклеточного потенциала, а также свойство этих ТПСП возникать при любых способах активации пирамидных нейронов гиппокампа, то не исключено, что и они участвуют в генерации суммарных экстраклеточных потенциалов θ-ритма гиппокампа. По-видимому, паузы между вспышками импульсной активности гиппокампальных нейронов, которые наблюдаются на фоне θ-ритма, вызваны не просто отсутствием возбудительных влияний, а активным торможением импульсной активности. В начале каждой паузы торможение может быть связано с католической депрессией в результате глубокой деполяризации, вызванной системой возвратной активации, а в поздней части паузы торможение может быть результатом гиперполяризации, вызываемой системой возвратного торможения.
Роль системы коллатерального торможения в гиппокампе заключается не только в том, что тормозные синапсы могут принимать участие в электрогенезе суммарного θ-ритма. Она в значительной степени определяет также временные характеристики ответов гиппокампальных нейронов на залпы импульсов из перегородки, и, возможно, именно система возвратного торможения является причиной того, что нейроны гиппокампа легко вовлекаются в генерацию θ-ритма при приходе ритмических залпов импульсов из перегородки. Процесс, облегчающий вовлечение нейронов гиппокампа в ритмическую активность, может заключаться в следующем.
Длительность возвратного постсинаптического торможения довольно велика — часто более 100 мс. По окончании периода торможения, вызванного активацией нейронов гиппокампа, можно наблюдать явление «отдачи»: после окончания гиперполяризации внутриклеточный потенциал не остается на уровне потенциала покоя, а переходит через уровень покоя в сторону деполяризации, сопровождающейся восстановлением импульсной активности нейрона. Это явление вначале было отмечено при экстраклеточной регистрации импульсной активности Грином и др. (Green at al., 1961), а затем — при непосредственной регистрации внутриклеточного потенциала пирамидных нейронов гиппокампа — Спенсером и Кэнделом (Spencer, Kandel, 1962). В результате этого повторная активация пирамидных нейронов гиппокампа с интервалом, равным длительности возвратного ТПСП или несколько большим, окажется значительно более эффективной, чем одиночная активация: повторное раздражение совпадает с «деполяризацией отдачи», снизившей порог активации нейронов. Но длительность возвратных ТПСП (100—200 мс) весьма близка к периоду θ-ритма, т. е. к длительности интервалов между ритмическими импульсами, посылаемыми в гиппокамп нейронами перегородки во время стресс-активности. Поэтому реакция нейронов гиппокампа на каждый залп импульсов из перегородки будет облегчаться «деполяризацией отдачи» от предшествующего залпа импульсов, и эффект окажется значительно сильнее, чем под влиянием таких же по интенсивности, но одиночных залпов импульсов (Spencer, Kandel, 1962).
Итак, обобщая сказанное, можно следующим образом представить себе последовательность процессов в различных структурах мозга во время θ-ритма. Активность нейронов перегородки, поддерживаемая импульсами от активирующих структур мозга, имеет тенденцию к группированию, — возникают залпы импульсных разрядов, следующие с частотой 4— 7 в 1 с. Эти ритмические залпы импульсов вызывают ответные ритмические реакции в ряде структур мозга, наиболее сильные — в гиппокамповой коре. Ответы пирамидных нейронов гиппокамповой коры на залпы импульсов из перегородки первично вызываются возбуждающими синапсами на базальных частях апикальных дендритов или на телах этих нейронов. Возбуждающие синапсы создают деполяризацию пирамидных нейронов, на фоне которой возникают импульсные разряды, которые поддерживают и углубляют начавшуюся деполяризацию, возможно, за счет возбуждающих возвратных связей. В результате этого деполяризация оказывается очень глубокой, что приводит к католической инактивации импульсных разрядов. После прекращения импульсных разрядов пирамидных нейронов деполяризация спадает или замещается гиперполяризацией. Эта гиперполяризация представляет собой ТПСП, вызванный посредством системы возвратного торможения импульсными разрядами пирамидных нейронов. Система возвратного торможения включает коллатерали аксонов пирамидных нейронов гиппокампа и вставочные тормозные нейроны — корзинчатые клетки гиппокампа. По окончании ТПСП наблюдается явление «отдачи» — вторичная деполяризация, облегчающая реакции нейронов на приходящие в это время импульсы из перегородки. Последнее явление значительно облегчает вовлечение нейронов гиппокампа в ритмическую активность. Медленным колебаниям внутриклеточного потенциала пирамидных нейронов гиппокампа (деполяризация и последующая гиперполяризация) по временным характеристикам и ряду свойств соответствуют суммарные электрические колебания θ-ритма, регистрируемые от гиппокампа. Отсюда можно заключить, что суммарные колебания обусловлены теми же синаптическими токами, которые вызывают колебания внутриклеточного потенциала.
Поскольку θ-ритм легче вызывается электрическим раздражением ретикулярной формации, она должна участвовать в организации влияний, вызывающих его. На большую роль ретикулярной формации ствола в возникновении θ-ритма гиппокампа у кошек указывают данные Эдди и сотрудников (Eddy et al,, 1971).
У кролика высокочастотная стимуляция среднемозговой ретикулярной формации обычно активирует гиппокампальный θ-ритм, а по мере нарастания силы стимуляции подавляет его. Стимуляция небольшой ограниченной области среднего мозга, по данным Тории (Torii, 1961), оказывает только угнетающее влияние на θ-ритм. Блокирующий эффект разрушения перегородки связывают с ключевым положением, которое эта структура занимает на ретикуло-гиппокампальных путях (Petsche, Stumpf, 1962). Это подтверждают уже упомянутые данные о характере разрядов нейронов перегородки.
Петче и Штумпф обращают внимание на возможную роль в осуществлении θ-ритма анатомической системы связей, описанной Наута (Nauta, 1958). Эта система включает двухсторонние связи структур обонятельного мозга с некоторыми отделами ствола, в частности с центральным серым веществом и ядрами покрышки, которые именуются лимбическими полями среднего мозга. Помимо прямой проекции волокон гиппокампа в ретикулярные отделы среднего мозга обнаружено существование путей, имеющих синаптические переключения в разных участках переднего и промежуточного мозга, в перегородке, латеральных отделах гипоталамуса, маммилярных телах. Эта система связывает большое число неспецифических образований, многие из которых влияют на нормальное развитие θ-ритма. Разрушение ряда ядер таламуса, гипоталамуса и среднего мозга у кроликов и кошек устраняет θ-ритм из различных отделов мозга.
Экспериментальные данные относительно участия структур системы Наута в формировании θ-ритма оказались противоречивыми. В последних работах Пармеджиани и др. (Parmeggiani et аl., 1971) показано, что одной активности структур обонятельного мозга недостаточно для поддержания θ-ритма при отсутствии постоянных ритмических влияний из перегородки. Видимо, параметры θ-ритма определяются взаимодействием импульсации из перегородки с активностью, приходящей из обонятельных структур так называемого круга Папеца.
Особое значение в генезе θ-ритма придается гипоталамусу, который служит важной переключительной станцией в системе афферентных путей гиппокампа (Torii, 1961; Parmeggiani, 1967).
Интересные соображения о генезе θ-ритма высказали В. Ю. Ильюченок и Г. Н. Банников (1968). Используя вещества, вызывающие θ-ритм (антихолинестеразные и холиномиметические), они сделали вывод, что в лимбической системе имеется свой (мускаринэргический) механизм, ответственный за возникновение θ-ритма, в котором большую роль играют структуры перегородки и задней гипоталамической области. Однако они полагают, что упорядоченный ритм в РФ и затылочных областях коры есть следствие возбуждения ретикулярной активирующей системы, и этот ритм не зависит от θ-ритма гиппокампа и перегородки.
В нашей лаборатории И. И. Полетаевой (1968) в опытах на крысах и кроликах было показано, что локальное введение новокаина в перегородку ядра средней линии таламуса и центральное серое вещество среднего мозга вызывает временное выключение θ-ритма во всех отделах мозга. В последнее время получены интересные данные (Тараненко и др., 1971), показывающие, что и новая кора играет, видимо, роль в способности гиппокампа генерировать θ-ритм, двустороннее удаление неокортекса полностью устраняло спонтанный и вызванный θ-ритм в гиппокампе.
Все эти данные заставляют полагать, что для генерации θ-ритма необходимо участие многих отделов мозга, возможно, объединенных в некоторую функциональную систему, более широкую, чем лимбическая система.
Таким образом, если перегородка и является водителем θ-ритма, то важное, а часто определяющее значение для проявления его играют многие структуры мозга, оказывающие влияние как на водитель ритма, так и непосредственно на структуры мозга, его генерирующие.
Определенная роль всех этих структур мозга в генерации θ-ритма, связь θ-ритма с реакцией пробуждения, ориентировочной реакцией и поведением, заставляют предполагать, что система структур, принимающих участие в генерации этого ритма, имеет важное значение для формирования целостной реакции организма.
Существуют, однако, мнения, ограничивающие роль перегородки в генерации θ-ритма в гиппокампе. Еще Дуглас и Прибрам (Douglas, Pribram, 1966) предположили, что есть два раздельных входа в гиппокамп: один из них «предназначен» для общего его возбуждения, другой— для ввода сенсорной информации.
Элазар и Эйди (Elazar, Adey, 1967) нашли, что на ранних стадиях обучения наблюдается θ-ритм с частотой 4—5 Гц, а по мере обучения она сдвигается в область 6—7 Гц. Были получены данные (Eddy et al., 1970), что эти два диапазона частот θ-ритма связаны с функционально различными нервными процессами и возникают в гиппокампе при активации различных механизмов. Так, частоту θ-ритма 4—5 Гц авторы связывают с ретикулярной формацией ствола и перегородкой, в то время как частоты 6—7 Гц со специфической информацией, приходящей к гиппокампу из других источников: субталамуса, миндалины, опосредовано через специфические пути и из энторинальной коры. Это подтверждается экспериментальными данными (Eddy et al., 1970), показывающими, что разрушение перегородки не угнетает в гиппокампе θ-ритма — 6—7 Гц. Значит, последняя лишь частично вовлечена в генерацию θ-ритма. С точки зрения этих авторов, гиппокамп представляется как. компаратор в вычислительной машине, получающий входы из двух источников и сравнивающий сходство и различия между ними.
Подчеркнем и другую сторону, регулирующую роль структур, генерирующих θ-ритм, на возбудимость отдельных структур мозга, Начало таким исследованиям положено Пармеджиаии и Рабини (Parmeggiani, 1962; Parmеggiani, Rabini, 1964), которые в остром опыте отметили изменение первичных ответов зрительной и слуховой зон коры и вызванных потенциалов теменно-центральной коры кошки (падение амплитуды) при возникновении θ-ритма на болевое раздражение и показали, что эти изменения достоверно ослабевают после выключения θ-ритма коагуляцией перегородки. Появление θ-ритма в гиппокампе кошки в ответ на раздражение промежуточного и среднего мозга (Jokota, Fujimori, 1964) сопровождается облегчением спинномозговых рефлексов, кожно-гальванической реакцией и подъемом кровяного давления.
Этот вопрос был специально исследован И. И. Полетаевой (1968). Она выяснила, что инактивация различных структур, генерирующих θ-ритм, оказывает облегчающее влияние на ретикулярную формацию среднего мозга, вызывает падение возбудимости гиппокампа и ослабление стабильности ряда корковых функций. Видимо, активное состояние структур, генерирующих θ-ритм, оказывает «притормаживающее влияние» на ретикулярную формацию среднего мозга и стабилизирующее и некоторое активирующее влияние на реакции новой коры, или, иными словами, они, возможно, обеспечивают уровень функционального состояния мозга при резких изменениях внешней среды.
В последние годы (Komisaruk, 1970) при изучении реакции исследовательского принюхивания у крыс было показано, что в момент этой реакции вибриссы подергиваются взад и вперед с частотой 7 в 1 с. Часто наблюдается прямая корреляция между каждым подергиванием вибрисс и каждой волной θ-ритма в лимбико-гипоталамической системе и соседних отделах мозга. Была выдвинута гипотеза, что лимбико-гипоталамическая система образует пульсирующий нейрональный выход, синхронный с θ-ритмом, который модулирует разряды мотонейронов, вовлеченных в ритмическое исследовательское принюхивание, через гипоталамические эфферентные пути к нижним отделам ствола и к спинному мозгу.
Фармакологическими исследованиями установлено, что в генерации θ-ритма принимают участие как адренореактивные (связанные преимущественно с формированием отрицательной реакции организма), так и холино- и серотонинореактивные системы (Ильюченок, 1965).
В последние годы (Ильюченок, 1971) было показано, что появление θ-ритма в гиппокампе при действии антихолинэстеразных и холиномиметических веществ есть следствие изменения активности М-холинэргического механизма лимбической системы, большую роль в котором играют холинэргические механизмы заднего гипоталамуса и перегородки.
То же показано (Пастухов и др., 1971) для лимбической системы и относительно адренэргических механизмов, которые при определенных условиях могут участвовать в активирующей деятельности независимо от адренэргических механизмов восходящей ретикулокорковой активирующей системы ствола мозга. Поэтому пиридрол и фенамин вызывают θ-ритм в гиппокампе при полном отделении ретикулярных структур ствола мозга. Это, конечно, не отрицает того, что в нормальных условиях целого мозга θ-ритм в значительной степени определяется системой ретикулярная формация — перегородка.
В действии веществ разных медиаторных групп выделяется ряд особенностей. Эзерин, например, вызывает четкий ритм, частота которого растет с повышением дозы, но не превышает 7 в 1 с, амфетамин вызывает синхронизацию ЭЭГ с частотой 4—5 в 1 с, независимо от дозы (Salier, Stumpf, 1957). По данным Петче и др. (Petsche et al., 1965), возбуждение структур, участвующих в генерации θ-ритма, может выражаться, во-первых, в увеличении числа нейронов перегородки, разряжающихся в ритме θ-волн с одновременным усилением синхронизации суммарной ЭЭГ (тип действия амфетамина), во-вторых, в увеличении частоты разрядов клеток перегородки (тип действия эзерина). В основе второго эффекта лежит изменение режима разрядов нейронов. Оба вещества усиливают приток импульсации из нижележащих отделов мозга, хотя эзерин, возможно, влияет и непосредственно на клетки перегородки.
Десинхронизация как реакция на раздражитель отмечается и в реакциях гиппокампа. Так, при интенсивном раздражении седалищного нерва (Green et al., 1960) реакция пробуждения в гиппокампе начинается с короткого периода десинхронизации, на смену которой приходит θ-ритм, Корацца и Пармеджнани (Corazza, Permeggiani, 1961), описавшие подобный эффект в опытах на кошках, предполагают, что в норме существуют два типа воздействия на гиппокамп — синхронизирующее и десинхронизирующее. Подтверждением могут служить уже известные факты, свидетельствующие, что реакция пробуждения гиппокампа в виде десинхронизации становится обычным явлением после разрушения перегородки, так же как и в ответ на высокочастотное раздражение перегородки. Введение небольших доз барбитуратов вызывает появление в гиппокампе медленных волн, сменяющихся десинхронизацией в ответ на любое внешнее раздражение (Weiss, Bolidanesku, 1969, 1963). В препаратах cervean isole (Kawamura et al., 1961) электрическая активность гиппокампа носила черты непрерывной активации, в то время как в новой коре преобладали вспышки веретен. На основании экспериментов с коагуляцией разных участков мозга авторы пришли к выводу, что основной активирующей системой для неокортекса является ретикулярная активирующая система, а для архи- и палеокортекса — гипоталамическая система активации.
Десинхронизацию в гиппокампе как реакцию на сверхпороговое раздражение гипоталамуса описал Граштиан с сотрудниками (Grastyan et al., 1966). На основании своих данных Штумпф (Stumpf, 1965) также предполагает, что синхронизирующая и десинхронизирующая системы пространственно различны и внешние воздействия (коагуляция, фармакологические, вещества) влияют на них в неодинаковой степени.
В онтогенезе кролика θ-ритм формируется к концу второй недели жизни (Gogolak et al., 1963; Никитина, Юсова, 1965). До этого времени гиппокамп реагирует на раздражение реакцией десинхронизации. Видимо, десинхронизирующая система формируется раньше, чем синхронизирующая.
Изложенные данные о доли структур мозга в генерации θ-ритма заставляют предполагать, что эти структуры должны играть важную роль и в формировании поведения животного.
Работа Грина и Ардуини (1954) в другие исследования показали, что электролитическое разрушение перегородки у крыс и кроликов, ведущее к исчезновению θ-ритма, делает животных необычно пугливыми и настороженными, повышает их чувствительность к раздражениям. Брейди в Наута (Brady, Nauta, 1953, 1955), а вслед за ними еще ряд исследователей наблюдали, что разрушение перегородки делает ручных крыс агрессивными, уменьшает величину отрицательной условной реакции. Отмечено, однако, что эти явления постепенно исчезают, причем независимо от приручения животных. Разрушение перегородки у кошек усиливает спонтанную двигательную активность, поисковую и ориентировочную реакции, подавить которые можно лишь введением больших доз хлорпромазина и барбитуратов. Еллен и др. (Ellen et al., 1962, 1964) обнаружили, что разрушение перегородки ослабляет процесс внутреннего торможения у крыс, а Цопф и Граштиан (Czopf, Grastyan, 1963) описывают ухудшение запаздывающих рефлексов у кошек в результате этой операции. Локальная инъекция карбохолина в перегородку снижает, а атропин и адреналин повышают безусловную реакцию избегания у крыс (Grossman, 1964).
Необходимо, однако, осторожно подходить к вопросу о функциональном значении θ-ритма, так как данных об этом еще совершенно недостаточно.