➥ Основная статья: Тета-ритмы
Классическая / гиппокампальная модель возникновения тета-ритма
Гиппокампальная модель генерации тета-ритма основывается на экспериментальных данных, полученных на мозге различных млекопитающих. Делается допущение, что механизм генерации тета-ритма у человека аналогичен механизмам, описанным для мозга млекопитающих. Исследования у млекопитающих позволили сформулировать следующую концептуальную модель возникновения тета-активности.
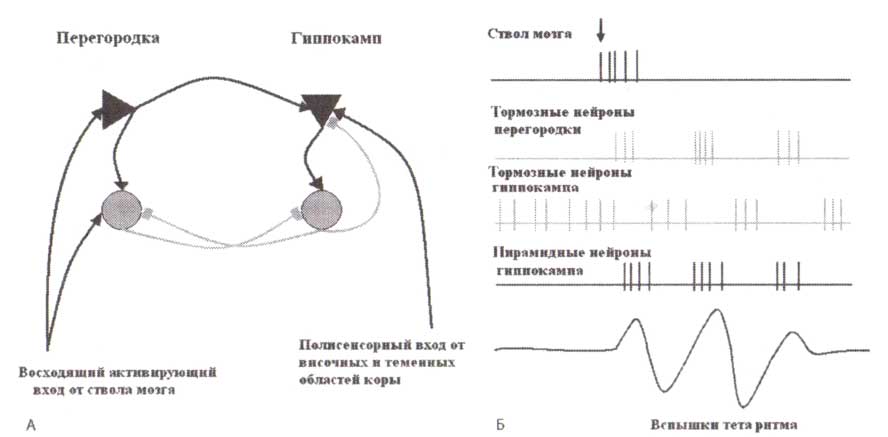
«Узловое» значение придается зоне перегородки в основании переднего мозга. Коллатерали нейронов афферентных систем при поступлении сенсорной информации любой модальности активируют нейронные элементы ствола мозга (обозначено стрелкой на рис. 1). От нейронов ретикулярной системы ствола, в частности норадренергических клеткок, расположенных в голубом пятне, серотонинергических клеткок — в ядрах шва, допаминергических клеткок — в передней части покрышки и компактной части черной субстанции, неритмические колебания поступают в перегородку. В состоянии спокойного бодрствования фоновая активность тормозных нейронов в гиппокампе подавляет гиппокампальные пирамидные клетки так, чтобы никакая сенсорная информация не кодировалась в след памяти. В случае же если появляется значимый стимул (например, эмоционально-значимый), этот стимул через активирующую систему ствола мозга увеличивает активность возбуждающих и тормозных нейронов перегородки. Тормозные нейроны перегородки снижают активность тормозных клеток гиппокампа. Это торможение активности тормозных клеток гиппокампа (растормаживание) удаляет фоновое торможение пирамидных клеток и в конечном счете способствует их работе. Такая последовательность нейронных событий запускает цикл гиппокампального тета-ритма. Исходя из описанной модели очевидно, что активация клеток ядер перегородки вызывает вспышки тета-ритма, а не длительную монотонную активность.
Далее активация пирамидных клеток гиппокампа тормозит тормозные клетки в ядрах перегородки, которые, в свою очередь, снимают торможение с гиппокампальных тормозных нейронов, а те далее тормозят пирамидные клетки. Так завершается цикл. Гиппокампальный тета-ритм зависит от силы входящих сигналов от нейронов перегородки: когда достигается некоторый критический уровень активации нейронов перегородки — возникает гиппокампальный тета-ритм. В гиппокампе животных существуют не один, а по крайней мере два типа ритмической тета-активности. Первый тип связан с произвольными движениями и не изменяется поддействием атропина или холинергического истощения. Второй тип тета-активности иногда возникает в состоянии неподвижности, снимается атропином и может спонтанно возникать при уретановой анестезии (Kramis и др., 1975).
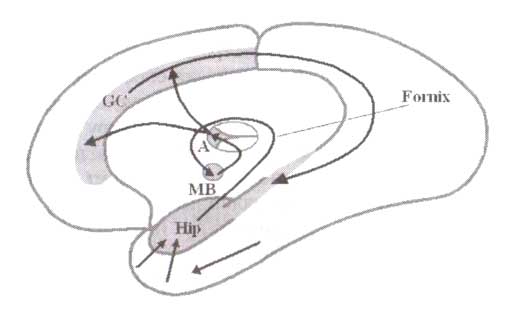
Гиппокампальный тета-ритм регистрируется не только во всех частях гиппокампа, но и «проводится» в структуры, связанные с гиппокампом анатомически (рис. 2). Эти структуры относятся к так называемой лимбической системе. Лимбическая система, отвечая за эмоции и участвуя в консолидации следов памяти, фактически представляет собой петлю циркуляции возбуждения — «круг Пейпеца». Информационные потоки в лимбической системе могут быть описаны следующим образом. Гиппокамп выступает как полисенсорный коллектор, принимая информацию с различных структур мозга. В гиппокампе происходит интеграция поступивших сигналов, и результаты обработки через пучок аксонов свода (fornix) поступают к мамиллярным телам. Мамиллярные тела, «сравнив» поступившую информацию с информацией от других источников, посылают «дискриминантный результат» к переднему ядру таламуса. Переднее ядро таламуса обрабатывает информацию и направляет сигнал к передней поясной извилине. Передняя поясная извилина по своим связям с гиппокампом замыкает «круг Пейпеца».
Гиппокампальная теория не дает исчерпывающих объяснений для всей ЭЭГ феноменологии. Так, накопилось уже достаточно данных, позволяющих обоснованно считать, что при определенных функциональных состояниях альфа-ритм может иметь частоту ниже общепринятых 8 Гц, занимая частоты, формально относящиеся к тета-диапазону. Выделяют так называемый «медленный вариант» альфа-ритма (slow alpha-variant rhythm), имеющий частоту 4—7 Гц, регистрируемый в задних отделах мозга. Таким образом, регистрируемая на ЭЭГ спонтанная активность частотой 4—8 Гц не всегда является собственно тета-ритмом.
Альфа-тета-континуум
Исследования механизмов генерации биоэлектрической активности мозга человека в настоящее время определили «таламическую» теорию генерации альфа-ритма как доминирующую. Эта гипотеза подразумевает наличие ключевого нейронального механизма: залповой активности у определенных таламокортикальных нейронов.
Было показано, что затылочный альфа-ритм возникает одновременно с ритмическими разрядами строго такой же частоты в нейронах LGN. Степень когерентности между активностью таламуса и активными областями коры всегда очень высока, что доказывает ключевую роль LGN в поддержании альфа-ритма.
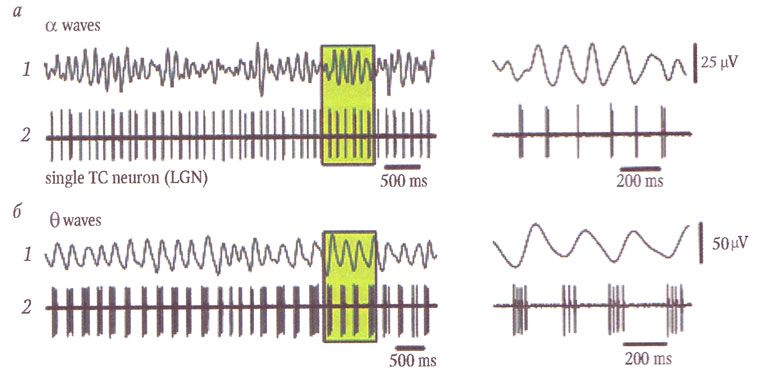
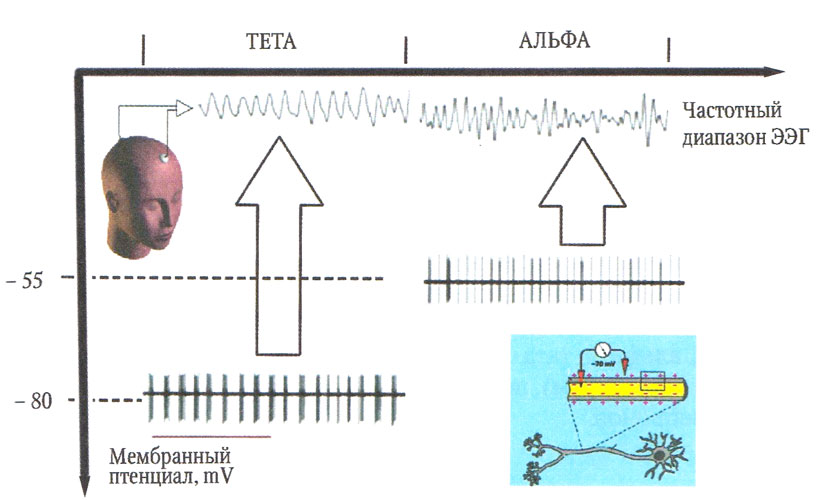
В работах Стюарта Хьюза (S. Hughes), которые уже можно признать классическими, показано, что отличительной особенностью данных таламокортикальных нейронов LGN является их способность генерировать повторяющиеся пачки разрядов, охватывающие не только диапазон альфа-ритма (8—13 Гц), но и разряды на более низких частотах — от 2 до 8 Гц (рис. 3). Это значит, что разрядная активность таламокортикальных нейронов охватывает весь диапазон тета- и альфа-частот. Фактором, определяющим частоту разрядной активности, является степень деполяризации нейронной мембраны. При «нормальном» уровне деполяризации (около -55 mV) генерируются двойные разряды с частотой, совпадающей с таламической и корковой альфа-активностью. При относительной гиперполяризации мембраны (ниже -55 mV) таламокортикальные нейроны переходят на генерацию пачек, содержащих от трех до пяти потенциалов действия (Hughes S. W., Crunelli V., 2005). Частота следования этих пачек варьирует от 1—3 до 7—8 Гц. Анализ взаимосвязи разрядной активности нейронов и суммарной ЭЭГ показал, что низкочастотным разрядам соответствуют вспышки тета-активности (2—7 Гц). Замещение происходит при снижении уровня бодрствования на ранних стадиях сна (рис. 4).
Следствием исследований разрядной активности таламических нейронов явился вывод о единстве механизмов, лежащих в основе генерации как альфа-ритма, так и тета-диапазона частот на ЭЭГ. На этом основании Стюарт Хьюз и Винченцо Крунелли (S. W. Hughes, V. Crunelli, 2005) ввели понятие электроэнцефалографического «альфа-тета-континуума». Согласно этой концепции альфа- и тета-диапазон частот на ЭЭГ рассматривается как результат консолидированной активности нейронов релейных таламических ядер и ретикулярного ядра таламуса. Предложенная концепция не противоречит теории гиппокампального механизма генерации тета-ритма. Благодаря своим обширным внутрицентральным связям активность нейронов древнего гиппокампа может рассматриваться как «двойник» залповой активности таламических нейронов.
Уровень деполяризации мембран таламокортикальных нейронов определяет активность глутаматергических проекций кортикальных нейронов, преимущественно лобной коры. Активация нейронов коры приводит к деполяризации таламических нейронов, что проявляется увеличением частоты вспышек и определяет формирование локальной электрической активности в альфа-диапазоне. И наоборот, дезактивация глутаматергических нейронов приводит к гиперполяризации, что переводит таламические нейроны на более низкую частоту генерации высокопороговых вспышек. Этот процесс лежит в основе генерации тета-ритма.
Роль нисходящих кортикоталамических проекций доказывается следующими экспериментами. Ритмические таламокортикальные колебания в диапазоне альфа-частот можно смоделировать in vitro на препарате среза латерального коленчатого тела — первичного релейного ядра зрительного анализатора (Hughes et al., 2004). В таком препарате таламуса корковая часть отсутствует, что делает невозможным обратные кортико-таламические взаимодействия. Они могут быть смоделированы фармакологической активацией глутаматных рецепторов таламических нейронов. Внутриклеточная регистрация активности таламокортикальных нейронов препарата среза таламуса показывает, что вспышки пейсмекерной активности происходят периодично в широком диапазоне частот от 4 до 13 Гц. Было обнаружено, что более сильная активация глутаматных рецепторов, имитирующая высокую активность нейронов фронтальной коры, ведет к появлению ритмов с альфа-частотами, тогда как уменьшение уровня активации вызывает замедление до частот тета-диапазона (4—8 Гц).
Таким образом, альфа- и тета-активность может рассматриваться как непрерывный спектр (континуум), поддерживаемый специфическими свойствами таламических нейронов.