Импульсная или спайковая активность (от англ, spike — острие) характерна для тел и аксонов нервных клеток и связана с бездекрементной (скачкообразной) передачей возбуждения от одной нервной клетки к другой, от рецепторов к центральным отделам нервной системы или от центральной нервной системы к исполнительным органам.
Импульсная активность нейронов возникает в результате достижения мембраной нейрона критического уровня деполяризации, при котором наступает электрический «пробой» мембраны и начинается самоподдерживающийся процесс распространения возбуждения в нервном волокне, который называется потенциал действия.
⚠️ Нейронный импульс, спайк и потенциал действия являются синонимичными понятиями. Не путать с ЭЭГ спайками.
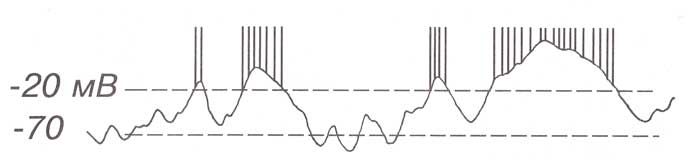
При внутриклеточной регистрации импульсная активность нейрона имеет вид высокоамплитудных, коротких, быстрых положительных пиков, которые часто следуют друг за другом. Такая серийная импульсная активность называется вспышкой.
Характерными особенностями нейронных спайков являются их высокая амплитуда (порядка 50-125 мВ), небольшая длительность (порядка 1-2 мс), приуроченность их возникновения к достаточно строго ограниченному электрическому состоянию мембраны нейрона (критический уровень деполяризации) и относительная стабильность амплитуды спайка для данного нейрона (закон «все или ничего»).
Кальциевый спайк
Кальциевый спайк — это восстановительная деполяризация таламокортикальных клеток, которая следует за сильной гиперполяризацией нейронов. Деполяризация обеспечивается кратковременным низкопороговым Са2+ -током (I1), который инактивируется, когда нейрон деполяризован, и деинактивируется при гиперполяризации.
Отличие импульсной активности нейронов от постсинаптических потенциалов
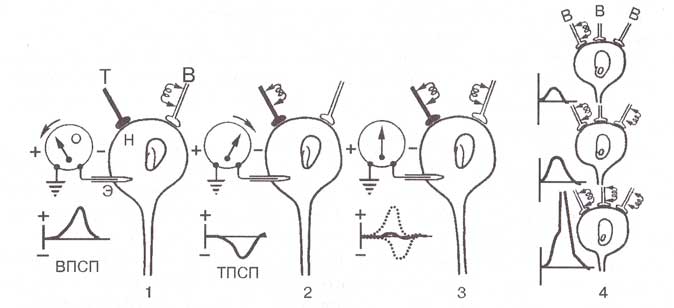
Постсинаптические потенциалы (ПСП) – это градуальные электрические реакции, присущие в основном дендритам в соме нейрона, возникающие в ответ на приход к нейрону электрических импульсов по афферентным путям от других нервных клеток.
В зависимости от активности возбуждающих или тормозящих синапсов соответственно различают возбуждающие постсинаптические потенциалы (ВПСП) и тормозные постсинаптические потенциалы (ТПСП). ВПСП проявляется положительным отклонением внутриклеточного потенциала, а ТПСП — отрицательным, что соответственно обозначается как деполяризация и гиперполяризация. Эти потенциалы отличаются локальностью, декрементным распространением на очень короткие расстояния по соседним участкам дендритов и сомы, сравнительно малой амплитудой (от единиц до 20-40 мВ), большой длительностью (до 20-50 мс). В отличие от спайка, ПСП возникают в большинстве случаев независимо от уровня поляризации мембраны и имеют различную амплитуду в зависимости от объема афферентной посылки, пришедшей к нейрону и его дендритам. Все эти свойства обеспечивают возможность суммации градуальных потенциалов во времени и пространстве, отображающей интегративную деятельность определенного нейрона12. Именно процессы суммации ТПСП и ВПСП определяют уровень деполяризации нейрона и, соответственно, вероятность генерации нейроном спайка, т.е. передачи накопленной информации другим нейронам (рис. 1).
Как видно, оба эти процесса оказываются тесно связанными: если уровень импульсной “бомбардировки”, обусловленной приходом спайков по афферентным волокнам к нейрону, определяет колебания мембранного потенциала, то уровень мембранного потенциала (градуальные реакции) в свою очередь обусловливает вероятность генерации спайка данным нейроном (рис. 2).
Как следует из изложенного выше, импульсная активность нейрона представляет собой значительно более редкое событие, чем градуальные колебания соматодендритного потенциала. Приблизительное соотношение между временным распределением этих событий можно получить из сопоставления следующих цифр: спайки генерируются нейронами мозга со средней частотой 10 в секунду; в то же время по каждому из синаптических окончаний к дендритам и соме притекает соответственно в среднем 10 синаптических воздействий за секунду. Если учесть, что на поверхности дендритов и сомы одного коркового нейрона могут оканчиваться до нескольких сотен и тысяч синапсов, то объем синаптической бомбардировки одного нейрона, а соответственно и градуальных реакций, составит несколько сотен или тысяч за секунду. Отсюда соотношение между частотой спайковой и градуальной реакции одного нейрона составляет 1-3 порядка. Относительная редкость спайковой активности, кратковременность импульсов, приводящая к их быстрому затуханию из-за большой электрической емкости коры, определяют отсутствие значительного вклада в суммарную ЭЭГ со стороны спайковой нейронной активности.
Связь спайковой активности и ЭЭГ
Таким образом, электрическая активность мозга отображает градуальные колебания соматодендритных потенциалов, соответствующих ВПСП и ТПСП. Связь ЭЭГ с элементарными электрическими процессами на уровне нейронов нелинейная. Наиболее адекватной в настоящее время представляется концепция статистического отображения активности множественных нейронных потенциалов в суммарной ЭЭГ. Она предполагает, что ЭЭГ является результатом сложной суммации электрических импульсов многих нейронов, работающих в значительной степени независимо. Отклонения от случайного распределения событий в этой модели будут зависеть от функционального состояния мозга (сон, бодрствование) и от характера процессов, вызывающих элементарные потенциалы (спонтанная или вызванная активность).
В случае значительной временной синхронизации активности нейронов, как это отмечается при некоторых функциональных состояниях мозга или при поступлении на корковые нейроны высокосинхронизированной посылки от афферентного раздражителя, будет наблюдаться значительное отклонение от случайного распределения. Это может реализоваться в повышении амплитуды суммарных потенциалов и увеличении когерентности между элементарными и суммарными процессами34.
Как показано выше, электрическая активность отдельных нервных клеток отражает их функциональную активность по переработке и передаче информации. Отсюда можно сделать заключение, что суммарная ЭЭГ также в преформированном виде отражает функциональную активность, но уже не отдельных нервных клеток, а их громадных популяций, т.е., иначе говоря, функциональную активность мозга. Это положение, получившее многочисленные неоспоримые доказательства, представляется исключительно важным для анализа ЭЭГ, поскольку дает ключ к пониманию того, какие системы мозга определяют внешний вид и внутреннюю организацию ЭЭГ.
Footnotes
- Костюк П.Г., Шаповалов А.И. Электрофизиология нейрона. В кн.: Современные проблемы электрофизиологических исследований нервной системы. М.: Медицина, 1964, с. 31-50.
- Eccles J.C. The Physiology of synapses. — Berlin-Gottingen-Heidelberg: Springer-Verlag, 1964.
- Speckmann E.J. and Walden J. Mechanisms Underlying the Generation of Cortical Field Potentials /Acta Otolaryngol (Stockh) 1991; Suppl. 491, p. 17—24.
- Zschocke St. Vom Neuron zum EEG: Die Entstehungsmechanismen des EEG. I. Die Potenialquellen des I IX i / EEG-Labor, 1991, 13, s. 43—57.