Одним из характерных видов суммарной электрической активности, регистрируемой в ряде структур обонятельного анализатора животных и человека, являются высокоамплитудные вызванные волны (ВВ), имеющие вид веретена, которые возникают в ответ на адекватную стимуляцию обонятельной выстилки. Частота ВВ в веретене отличается у животных разных классов и зависит от температуры тела (Гусельников, 1956; Takagi, Shibuya, 1960). Линейная зависимость частоты ВВ от температуры тела была найдена у каймана, у двоякодышащих рыб и у кролика (Huggins et al., 1968; Pitkonen, Sarajas, 1968; Dupe, Gobet, 1969). Однако у всех видов животных, кроме рыб, частота волн всегда падает к концу веретена. Так, для лягушки показано (Гусельникова, Гусельников, 1975), что если длительность одного колебания в начале веретена равна 40— 50 мс, то в конце она возрастает до 140—160 мс. Меняется и форма колебаний: у начальных колебаний наблюдается короткая фаза нарастания негативности и более длительная фаза развития позитивности, а у последних— соотношение длительности фаз противоположное. У рыб частота волн к концу веретена может иногда увеличиваться.
Спонтанные вспышки ВВ появляются в ритме дыхания на вдохе и в ряде случаев на выдохе (Ueki, Domino, 1961). В хронических условиях опыта в обонятельной луковице (ОД) кошки ВВ возникают на различного рода сенсорные раздражения и раздражение ретикулярной формации. Поэтому была выдвинута гипотеза о том, что эта активность не столько связана с активностью обонятельных рецепторов, сколько с состоянием бодрствования в связи с активирующим влиянием ретикулярной формации на ОЛ и другие обонятельные центры. Однако последующие опыты показали (Domino, Ueki, 1960; Goult, Coustau, 1965), что вспышки веретен возникают только в том случае, если одновременно с центральными факторами имеет место раздражение воздухом обонятельных рецепторов. Поэтому предполагается, что центральные влияния действуют опосредованно через изменение дыхательного акта. Однако Пагано (Pagano, 1966) нашел, что одно только раздражение ретикулярной формации, хотя и не вызывало ВВ в ОЛ, но заметно влияло на них. Увеличение потока воздуха над обонятельной выстилкой так же вызывало увеличение ВВ. Таким образом, ВВ изменяются по амплитуде и длительности в зависимости от двух факторов: величины потока воздуха, проходящего через носовую полость, и центрифугальных влияний. Необходимость возбуждения элементов обонятельной выстилки для возникновения ВВ доказывается также опытами с орошением носовой полости новокаином; ВВ в ОЛ в этом случае исчезали. На кролике это было показано Л. А. Новиковой и Г. Я. Хволес (1953), и нами (Гусельников, 1956) — на черепахе. Однако ВВ не генерируются на уровне обонятельной выстилки, они генерируются элементами самой ОЛ. так как сохраняются в ней после полного ее отделения от остальных структур мозга, что показал еще Баумгартен с сотрудниками (Baumgarten et al., 1962). Это также подтвердилось в опытах нашей лаборатории на разных видах животных (Воронков, 1969; Энговатов, 1971; Гусельникова, Кипор, 1973, и др.).
Остается неизвестным, возбуждение каких рецепторов приводит к появлению ВВ; возможно, определенную роль в этом процессе играет механический фактор. Известно, что в эпителии обонятельного органа, помимо ольфакторных, имеются тригеминальные рецепторы и рецепторы якобсонова органа (Винников, Титова, 1957; Tucker, 1963, и др.). Обонятельный эпителий состоит из обонятельных, опорных и базальных клеток. В якобсоновом органе эпителий имеет то же строение, что и в обонятельной области основной полости носа; нервные волокна идут вместе с обонятельным нервом и оканчиваются в добавочной обонятельной луковице (Винников, Титова, 1957). Тригеминальная иннервация хорошо выражена в неольфакторных участках эпителия носовой полости (Moulton, 1965). Считается, что окончания волокон тройничного нерва сами выполняют роль рецепторов. Однако тройничный нерв не играет существенной роли в формировании ВВ, поскольку перерезка этого нерва, так же как и перерезка ножек мозга, не устраняет этих волн (Новикова, Хволес, 1953; Baumgarten et al., 1962; Маncia et al., 1962; Green et al., 1963).
Роль рецепторов якобсонова органа в генерации ВВ выяснена недостаточно. Восприимчивость этих рецепторов к механическим раздражениям (Adrian, 1954) и четкая реакция на обонятельные раздражители (Tucker, 1963) позволяют предположить, что рецепторы якобсонова органа участвуют в генерации волн. Однако аксоны этих рецепторов оканчиваются в добавочной ОЛ; между тем есть данные, свидетельствующие о том, что ВВ генерируются только элементами основной ОЛ (Adrian, 1950; Baumgarten et а!., 1962; С. Jamamoto, Т. Jamamoto, 1962).
Могут ли сами обонятельные рецепторы нести помимо обонятельной механорецепторную функцию, остается неясным, так же как неясно, присутствуют ли механорецепторы в обонятельном эпителии. Многие данные свидетельствуют о существенной роли механического фактора в возникновении обонятельных ВВ и обонятельного ощущения. Новиковой и Хволес (1953), например, удалось вызвать волны в ОЛ кролика при механическом раздражении обонятельного эпителия. Известно также, что для возникновения ВВ необходима стимуляция обонятельного эпителия током воздуха. Трахеотомия, блокада рецепторов новокаином и кокаином и перерезка обонятельного нерва приводят к исчезновению ВВ.
Долгое время оставалось неясным, какие причины способствуют возникновению ВВ: движение воздуха или какое-то пахучее вещество, которое всегда примешано к лабораторному воздуху. Некоторая ясность внесена и в этот вопрос. Так, Бейдлер (Beidler, 1958) показал, что скорость потока воздуха над обонятельным эпителием важна для возбуждения обонятельных рецепторов. Об этом свидетельствуют также данные других исследователей. Хагес и Мазуровский (Hughes, Mazurowski, 1962) нашли, что пропускание очищенного воздуха приводит к возникновению ВВ и ОЛ, Однако пороговая скорость потока для этого должна быть выше, чем в случае с добавлением пахучего вещества. Авторы полагают, что механическое раздражение рецепторов в естественных условиях может иметь место лишь при усиленном дыхании.
Уеки и Домино (Ueki, Domino, 1961) стимулировали обонятельный эпителий собак и обезьян потоком очищенного воздуха или газа (кислорода, азота, двуокиси углерода). Скорость и объем потока были близки к естественным, Амплитуда ВВ возрастала при увеличении в потоке доли очищенного газа, т. е. при уменьшении концентрации пахучего вещества. Уеки и Домино пришли к выводу, что механорецепторы существуют в обонятельной полости носа и ответственны за возникновение ВВ. Однако этому явлению можно дать и другое объяснение. Так, некоторые авторы полагают, что роль механического фактора заключается главным образом в переносе частиц пахучего вещества к рецепторам или в изменении турбулентных свойств потока. В конечном итоге это ведет к осмотическому раздражению обонятельных рецепторов (Green et al., 1963). В опытах на человеке Бокка и сотрудники (Восса et al., 1965) показали значительную роль механического фактора в формировании обонятельного ощущения.
Все приведенные факты, несомненно, свидетельствуют о важности механического фактора для возникновения ВВ. Однако основное значение в этом процессе, видимо, принадлежит ольфакторной функции обонятельных рецепторов, определяющей качественную и количественную характеристику ВВ. Исследования, проведенные Эдрианом (Adrian, 1942), Такаги с сотрудниками (Takagi et al., 1960), Хагесом и Мазуровским (Hughes, Maziirowski, 1962) и др. дают основание для такого предположения. В этих работах показано, что некоторые пахучие вещества могут угнетать вызванные волны. К такому же эффекту приводит увеличение концентрации вещества. Каждое пахучее вещество имеет свою характеристику частотного максимума в картине частотного спектра вызванных волн (Hughes, Mazurowski, 1962, 1964).
Происхождение и механизмы вызванных волн
различных точек по глубине луковицы
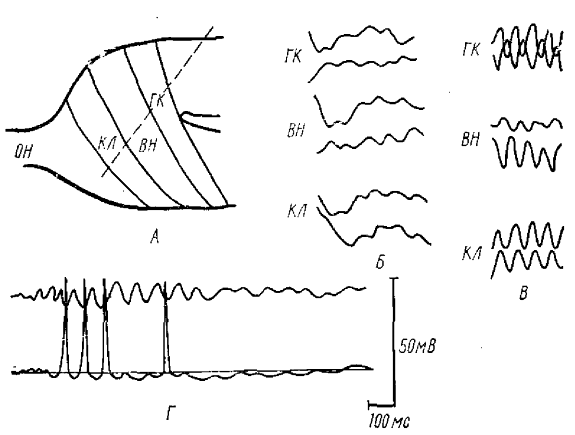
А—схематический разрез ОЛ лягушки с изображением основных слоев, Пунктир — направление движения отводящего электрода. Г — соотношение фаз ВВ (верхняя запись) с внутриклеточно регистрируемой активностью ВН (нижняя запись). Отрицательность вниз (по Гусельниковой и Гусельникову, 1975). Слои — ОН — волокон обонятельного нерва, КЛ — клубочков, ВН — вторичных нейронов, ГН — гранулярных клеток. На Б и В нижняя кривая— запись от ростральной части ОЛ в одной и той же точке в течение всего опыта; верхняя — ответ на различной глубине при погружении электрода
Механизм ВВ наиболее хорошо изучен у амфибий и частично у рептилий, тогда как у млекопитающих их генез остается неясным. Поскольку механизмы ВВ обонятельного анализатора у рыб, амфибий и рептилий были вскрыты в экспериментальных исследованиях, в основном в нашей лаборатории (Гусельникова, Гусельников, 1975; Энговатов, 1971; Гусельникова, Энговатов, 1973), то обратимся к фактическому материалу, ВВ регистрируются по всей поверхности ОЛ лягушки, однако наблюдается некоторое фазовое смещение волн в более каудальных точках относительно ростральных. При перемещении электрода по поверхности или погружении его вглубь ОЛ под углом (с целью прохождения всех слоев ОЛ) можно наблюдать инверсию ВВ близко к слою вторичных нейронов (ВН) (рис. 1, А, В). Характерным является то, что инверсия ВВ по глубине ОЛ происходит не через изолинию, т. е. не отмечается уровень, где амплитуда ВВ имела бы нулевое значение.
Точно такие же изменения претерпевает и ритмический разряд последействия (РРП) ОЛ, возникающий на ортодромное раздражение (рис. 1, Б). Эти факты свидетельствуют о сложной природе синаптических генераторов ВВ и о существенной роли ВН.
Тот факт, что амплитуда ВВ максимальна в районе обонятельных клубочков, говорит о том, что основной генератор ВВ расположен именно в этой ростральной части ОЛ. Изучая соотношение разряда ВН с фазой ВВ, мы обнаружили (рис. 1, Г), что они разряжаются на негативной их фазе.
У млекопитающих еще Эдриан (Adrian, 1950), изучая ВВ в ОЛ кролика, нашел, что они представляют собой следствие синхронного разряда большого количества ВН. Он предположил, что за генерацию ВВ ответственны дендриты митральных клеток (МК) ОЛ.
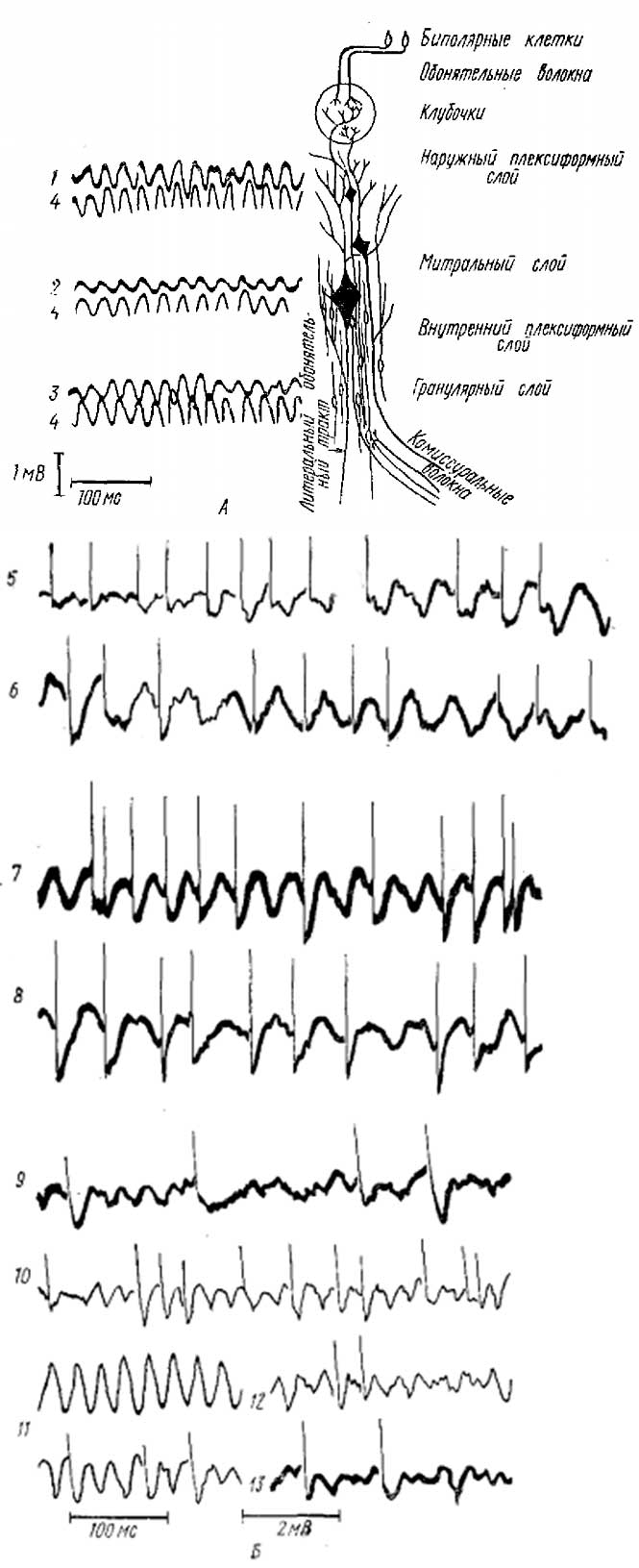
А — суммарная активность поверхности и глубоких слоев луковицы. 1—3 — запись от наружного плексиформного (1), митрального (2) и гранулярного (3) слоев (см. справа схему строения ОЛ); 4 —запись от контрольного электрода, постоянно расположенного на поверхности ОЛ около места введения погруженного электрода. Б—одновременная регистрация импульсной и суммарной активности ОЛ в слое митральных клеток (5, 6, 7), кисточковых клеток (7, 8), гранулярных клеток (9—13) (на записи 10 верхняя кривая активность, отводимая от поверхности ОЛ). Позитивность активных электродов соответствует отклонению вверх
Более подробный анализ ВВ в ОЛ кролика провели Баумгартен с сотрудниками (Baumgarten et al., 1962). Анализ распределения потенциалов в слоях обонятельной луковицы показывает, что ВВ обнаруживают характерную картину дипольного переворота (рис. 2, А): при погружении регистрирующего электрода в глубину ОЛ ритмические колебания, регистрируемые этим электродом, вначале остаются синфазными с колебаниями, регистрируемыми от поверхности, но при погружении регистрирующего электрода ниже слоя МК полярность колебаний меняется и они становятся противофазными по отношению к поверхностным потенциалам. Такая картина распределения потенциалов указывает на то, что основным генератором ВВ в ОЛ являются токи, направленные вдоль оси МК, которые вызывают колебания внутриклеточного потенциала тел этих клеток и соответствующие им колебания экстраклеточных потенциалов. С таким представлением согласуются данные о соотношении импульсной активности МК с ВВ: импульсные разряды, которые должны соответствовать деполяризационному изменению внутриклеточного потенциала, совпадают с негативно направленной фазой каждой волны ВВ в точке регистрации. Но одновременное исследование импульсной активности нейронов глубоких слоев ОЛ (гранулярных клеток— ГК), показывает, что на самом деле картина несколько более сложна. Разряды ГК наблюдаются на негативно направленной фазе волны, регистрируемой тем же микроэлектродом, что и импульсная активность (рис. 2, Б). Таким образом, поверхностные (МК и КК) и глубинные (ГК) клетки разряжаются в противофазе: разряды всех клеток соответствуют негативно направленной фазе ВВ в том же слое, где эти клетки находятся (а для поверхностных и глубоких слоев, как указывалось, ВВ противофазны). Эти результаты дают основание считать, что ВВ в глубоких слоях ОЛ являются не отражением (или не только отражением) активности МК, а генерируются также ГК. Следовательно, в ОЛ имеется, по крайней мере, две системы нейронов, которые являются согласованно действующими генераторами ритмической активности. Четкая корреляция активности отдельных нейронов с ВВ показывает, очевидно, что синхронность работы отдельных элементов во время генерации ритмической активности весьма высока. Данные, полученные нами на ОЛ амфибий, весьма сходны с данными, полученными на ОЛ млекопитающих.
Выдвигалось несколько гипотез о механизмах формирования ВВ и ОЛ, Так, например Оттосон (Ottoson, 1959) также полагал, что ВВ являются следствием синхронного возбуждения ВН за счет афферентного возбуждения. Однако эти авторы подчеркнув необходимость синхронизации активности нейронов ОЛ для ВВ, не объяснили ее происхождения.
Исследования, проведенные в нашей лаборатории, показали, что ВВ в ОЛ являются отражением локальных, синаптических процессов в нейронах ОЛ. Синаптические же механизмы генерации ВВ и ОЛ могут быть вскрыты только на основе тщательного внутриклеточного исследования течения процессов возбуждения и торможения. На млекопитающих таких работ не проводилось.
Этот вопрос был подвергнут анализу на ОЛ амфибии, рыб и рептилий с использованием внутриклеточной регистрации активности нейронов с одновременной регистрацией суммарной активности ОЛ при различных формах эксперимента. Прежде чем непосредственно перейти к механизмам вызванных волн ОЛ, необходимо рассмотреть синаптические процессы возбуждения и торможения в нервной сети ОЛ, их особенности и происхождение.
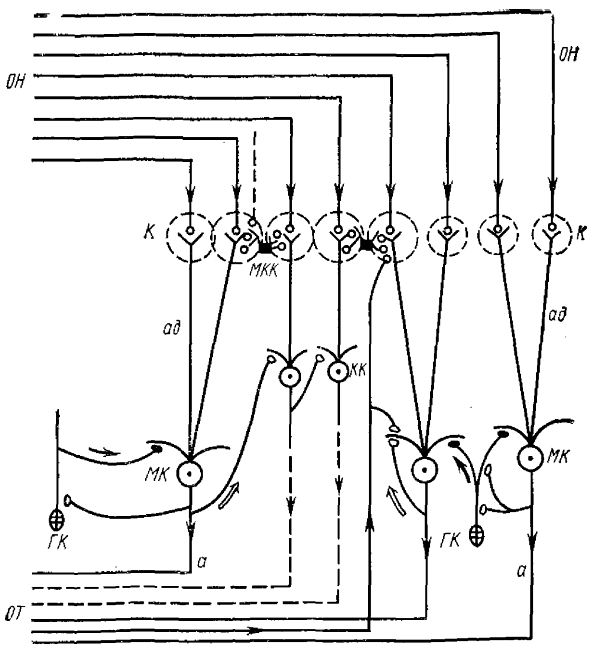
К — клубочки, МК — митральные, КК — кисточковые, МКК — межклубочковые и ГК — гранулярные клетки, ОН — волокна обонятельного нерва, ОГ — волокна обонятельного тракта, ад — апикальные дендриты вторичных нейронов, а—аксоны вторичных нейронов. Светлые стрелки и кружочки — направление возбуждающих влияний и возбуждающие синапсы; темные — направление тормозных влияний и тормозные синапсы
Для лучшего понимания этих процессов на рис. 3 дана принципиальная схема строения и связей ОЛ лягушки, построенная нами на основании литературных и собственных как морфологических, так и физиологических данных (Гусельникова, Гусельников, 1975). Мы рассмотрим более широко материал, связанный с генезом ВВ, преимущественно на ОЛ лягушки.

А — афферентный ВПСП, генерируемый им спайк и возвратный ВПСП во вторичном нейроне (1). Другой нейрон (2, 5). Вторичный нейрон без гиперполяризации (2), возвратный ВПСП генерирует спайк. Вторичный нейрон (а) — на фоне гиперполяризации возвратный спайк блокирован н возрос по амплитуде; б — возникновение возвратных ВПСП н спайков при разном способе возбуждения вторичного нейрона (4 и 6 — раздражение соответственно ипси- и контрлатерального обонятельного нерва, 5 — антидромное раздражение, 7— внутриклеточное раздражение нейрона); В — возникновение повторной деполяризации без предшествующего спайка во вторичном нейроне (8 — обычное возникновение возвратного ВПСП После афферентного спайка на ортодромное раздражение, 9 и 10 — один и тот же нейрон, 5 — спайк на ортодромное раздражение не развивается из-за рефрактерности. вызванной спонтанным спайком, 10-спайк на ортодромное раздражение блокирован гицерполяризующим током, проходящим через микроэлектрод); Г — Схема возвратного возбуждения в обонятельной луковице лягушки (ВН — вторичные нейроны, ад — апикальный дендрит, бд — базальный дендрит, а — аксоны). Светлые квадратики — возбуждающие синапсы
Примерно у 30% исследованных ВН в ответ на ортодромное раздражение (электрическая стимуляция обонятельного нерва) возникало деполяризационное колебание потенциала, на котором часто генерировался спайк (рис. 4). Проведенный анализ показал, что это афферентный ВПСП и развивается он на апикальных дендритах ВН в клубочках.
Во вторичных же нейронах на ОР регистрируется повторное деполяризационное колебание потенциала (рис. 4, А), которое резко отличается по своим свойствам от афферентного ВПСП. Это — возвратное возбуждение (или возвратный ВПСП). Возвратное возбуждение было обнаружено у 70 ВН из 144, причем оно характерно для КК ОЛ в 100% случаев, в то время как для МК только в 50% случаев. Какие можно привести доказательства, что повторное деполяризационное колебание является ВПСП и что оно возвратной природы?
Во-первых, оно увеличивается на фоне гиперполяризации нейрона. Оно не является также следовой деполяризацией, так как может возникать без первого спайка. Кроме того, как в МК, так и в КК этот ВПСП возникает при любом способе активации нейрона: ортодромном, антидромном раздражении, контрлатеральном и даже при прямом раздражении сомы нейрона током (см. рис. 5, Б). Провоцирование повторного ВПСП и спайка (а его, кроме того, часто провоцирует и спонтанно возникший спайк) любым способом типично именно для возвратного процесса, так как коллатерали аксонов должны активироваться независимо от способа возбуждения нейрона.
Поскольку повторный ВПСП (см. рис. 4) развивается после ТПСП, то возник вопрос — не является ли он результатом «отдачи», т. е. следствием постанодальной экзальтации нейрона, как это было показано для нейронов таламуса. Во-первых, необходимо отметить, что повторный ВПСП может возникать и без предшествующего ТПСП (см. рис. 4, Д). Во-вторых, специальный экспериментальный анализ показал, что повторные ВПСП и спайк действительно являются возвратными и что синапсы возвратного возбуждения расположены на базальных дендритах ВН, а само возвратное возбуждение осуществляется через коллатерали аксона (Гусельникова, Гусельников, 1975).
Хотя, как было нами показано, возвратное возбуждение и может быть локальным относительно данного нейрона, т. е. возможна возвратная связь на тот же самый нейрон; статистический вес такой связи невелик. Надо полагать, что возвратное возбуждение осуществляется в группе близлежащих нейронов перекрестными связями благодаря коллатералям аксонов ВН. Участие вставочного нейрона в цепи возвратного возбуждения не обязательно, так как никаких данных в пользу его существования мы не нашли (см. рис. 4, Г).
Возвратное возбуждение, видимо, играет большую роль в создании фонового спонтанного возбуждения ВН. В пользу этого свидетельствует ряд полученных нами фактов. Во-первых, гиперполяризующий ток, проходящий через микроэлектрод, блокирует спонтанный спайк, не оказывая обычно влияния на афферентный спайк (см. рис. 4). Во-вторых, спонтанный спайк часто вызывает возвратный спайк, что создает определенный узор спонтанной активности. И, наконец, во ВН иногда в ответ на ортодромное и часто на адекватное раздражения можно наблюдать длительную гиперполяризацию, которая не является ТПСП, так как на фоне искусственной гиперполяризации мембраны нейрона током она увеличивается по амплитуде. На основе этого мы рассматриваем ее как блок возбуждения, т. е, под влиянием определенных условий к нейрону может поступать меньший поток возбуждающих влияний через синапсы, расположенные на базальных дендритах, что поведет к повышению уровня поляризации мембраны нейрона. Доказательством того, что в этом случае основная роль принадлежит синапсам на базальных дендритах, служит тот факт, что эта медленная гиперполяризация значительно сильнее зависит от тока, проходящего через микроэлектрод, чем афферентный ВПСП.
Тормозные процессы в нейронной сети ОЛ лягушки и их соотношение с возбудительными процессами
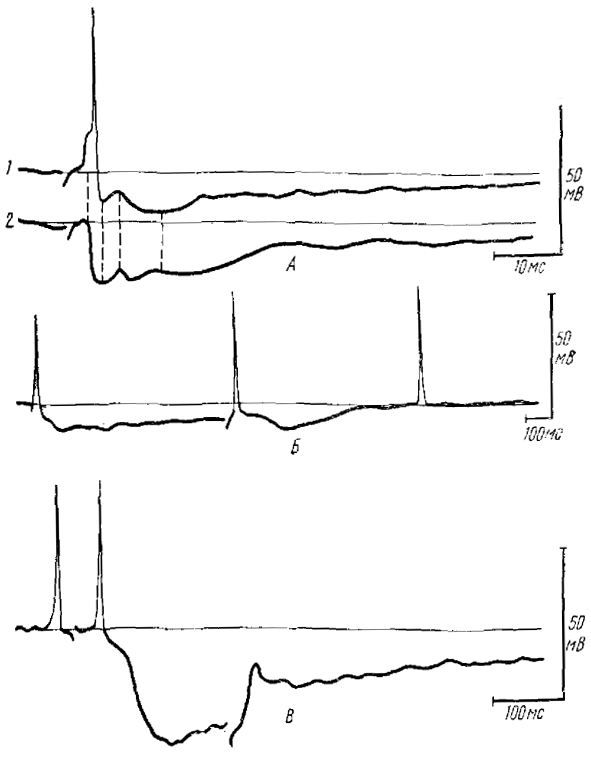
А — внутриклеточная регистрация одного и того же вторичного нейрона (/ и 2) при раздражении обонятельною нерва (начато ортодромного раздражения отмечено артефактом); 2 — наблюдается блокада генерации потенциала действия и уменьшение афферентного ВПСП за счет деполяризации нейрона через 20 мин после начала регистрации. Отчетливо выявляется латент-ность раннего ТПСП (первая пунктирная линия), максимальное развитие раннего ТПСП (вторая пунктирная линия) и вершина возвратною ВПСП (третья пунктирная линия) и повторное деполяризационное колебание нa фоне позднего ТПСП (четвертая пунктирная линия); Б — возникновение в МК двух фаз торможения — раннего и позднего (на повторное ортодромное раздражение, примененное на фоне позднего торможения, раннее торможение не возникает); В — на повторное ортодромное раздражение, нанесенное на фоне позднего торможения, афферентный ВПСП, не блокируемый ранним ТПСП, выявляется в полной мере. Раздражение на Б и В отмечено артефактом
Нам впервые удалось исследовать тормозные постсинаптические процессы во ВН ОЛ амфибий, показать их сложность и вскрыть генез (Гусельникова, Гусельников, 1975). На ортодромное и антидромное раздражения во ВН ОЛ лягушки сразу после афферентного ВПСП и спайка развивается гиперполяризация, которая сопровождается паузой в разряде нейрона (рис. 5).
Как правило, гиперполяризация, возникающая на ортодромное раздражение, является сложной по своему составу и отражает два последовательно развивающихся торможения: раннее и позднее. Как ранняя, так и поздняя фазы гиперполяризации являются ТПСП, так как они могут возникать без предшествующего спайка, понижают возбудимость нейрона, тормозят его разряд и на фоне искусственной гиперполяризации мембраны нейрона уменьшаются по амплитуде.
Однако ранний и поздний ТПСП отражают самостоятельные системы торможения, так как они могут появляться независимо друг от друга как в разных, так и в одном вторичном нейроне (рис. 5).
Латентный период раннего ТПСП равен примерно 10 мс, т. е. является очень коротким и в некоторых ВН не превышает латентного периода потенциала действия афферентного спайка. Длительность раннего ТПСП порядка 60—150 мс (вторичный ВПСП следует за ранним ТПСП) Латентный период позднего ТПСП относительно потенциала действия нейрона на ортодромное раздражение (ОР) —50—100 мс, а длительность его достигает 1—5 с.
Ранний ТПСП возникает на ортодромное раздражение, на антидромное не возникает или возникает с большим трудом. Он легко блокирует афферентный ВПСП и иногда спайк на ОР, Этот, а также ряд других полученных нами фактов позволили сделать вывод о близком расположении синапсов афферентного ВПСП и раннего ТПСП, т. е. ранний ТПСП генерируется на апикальных дендритах ВН.
Градуальные свойства раннего ТПСП выражены слабо, с увеличением силы ортодромного раздражения скачкообразно появляется новая волна ТПСП.
Если время возникновения раннего ТПСП совпадает с моментом развития в нейроне возвратного ВПСП, то вероятность возникновения раннего ТПСП значительно возрастает,— он может появляться в нейроне на тестирующий стимул (при парном раздражении), даже если не регистрировался на обусловливающий, При асфиксии, сезонных изменениях он либо не возникает, либо возникает очень редко. Эта свойства раннего ТПСП в значительной степени определяют, как мы увидим, и свойства ВВ, так как он является одним из главных составляющих волны веретена обонятельных ВВ.
Механизм генерации раннего и позднего торможения
Если синапсы раннего торможения расположены на апикальных дендритах ВН, то возникает вопрос, какие структуры образуют эти синапсы и какие структуры в ОЛ лягушки ответственны за генерацию раннего ТПСП?
Можно было полагать, что апикальные дендриты ВН получают тормозные влияния прямо от волокон обонятельного нерва, либо на пути генерации раннего ТПСП находится вставочный нейрон.
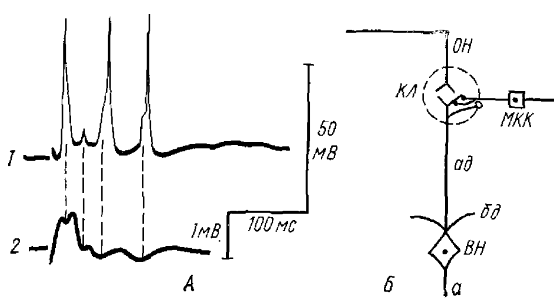
КЛ-клубочки, МКК — межклубочковые клетки, ВН — вторичный нейрон, ОН —волокна обонятельного нерва, ад — апикальный дендрит, бд — базальный дендрит, а — аксон. Белый кружок — возбуждающий синапс, томный — тормозный синапс
Исследования показали, что первое допущение не имеет места, тогда необходимо допустить наличие интернейрона в цепи раннего торможения. Этот вывод хорошо согласуется с морфологическими данными, согласно которым в клубочках имеют место межклубочковые клетки (МКК), которые по всем морфологическим данным можно считать тормозными. Мы, в свою очередь, располагаем физиологическими данными, которые подтверждают морфологические. Так, нам удалось показать, что потенциалы действия МКК соответствуют по времени позитивным фазам ВВ и ритмического разряда последействия в ОЛ, которые обычно отражают ТПСП во ВН (рис. 6, А: 8, Б).
Дальнейшие электрофизиологические и электронномикроскопические исследования (Гусельникова и сотрудники, 1971) показали, что активация МКК на ортодромное раздражение осуществляется через дендро-дендритные синапсы, образуемые дендритами ВН на дендритах МКК. Таким образом, возникновение раннего ТПСП можно представить следующим образом: в ответ на ортодромное раздражение на апикальных дендритах ВН возникает афферентный ВПСП, который генерирует дендритный спайк, возбуждающий через дендро-дендритный синапс. МКК, а последняя в свою очередь вызывает генерацию раннего ТПСП на тех же апикальных дендритах ВН (рис. 6, Б).
Позднее торможение возникает как на ортодромное, так и на антидромное раздражение, причем только в МК. В КК позднее торможение не возникает. Тот факт, что позднее торможение возникает в ОЛ при отделении последней от остального мозга, говорит о том, что оно генерируется структурами самой ОЛ.
Возникновение позднего торможения на ортодромное и антидромное раздражение (и даже лучше на антидромное) дало основание предполагать, что природа этого торможения возвратная. Этот вопрос был подвергнут специальному экспериментальному выяснению (Гусельникова, Гусельников, 1975), где возвратная природа позднего торможения была подтверждена.

ГК — гранулярные клетки, МК — митральные клетки. Остальные обозначения см. на рис. 6
Длительный латентный период позднего торможения и длительность самого торможения наводит на мысль об участии в этом процессе интернейрона. И действительно, нам удалось в гранулярном слое ОЛ зарегистрировать несколько ГК, которые обнаруживают свойства, типичные для интернейронов. В ответ на ортодромное и антидромное раздражение в этих клетках генерируется длительный топический ВПСП, на котором возникают спайки (рис. 7, А). Эти нейроны относятся к высокопороговым, а спонтанная активность у них либо вообще отсутствует, либо очень низкая (1 спайк в 10 мин).
Что касается локализации синапсов возвратного торможения, то они расположены вне сомы ВН, преимущественно на базальных дендритах. Это подкрепляется морфологическими данными, которые говорят о том, что дендриты ГК образуют синапсы главным образом на базальных дендритах МК и редко на их телах (рис. 7, Б).
Возвратное торможение является латеральным по своему характеру. Об этом говорит прежде всего то, что возвратный ТПСП может возникать без предшествующего спайка как на орто- так и на антидромное раздражение. Кроме того, спонтанные спайки не вызывают возвратный ТПСП, как не вызывает его и прямое раздражение нейрона током. Это говорит о том, что возвратная тормозная локальная связь (на тот же самый нейрон), по-видимому, не имеет места, т. е. возвратное торможение осуществляется за счет широких латеральных связей между нейронами.
Таким образом, в свете полученных данных о возбудительных и тормозных процессах в ОЛ лягушки находит объяснение ряд фактов, о которых мы упоминали, но не объясняли.
- Поскольку синапсы возвратного возбуждения и возвратного торможения расположены на базальных дендритах МК, объясняется тот факт, что возвратное возбуждение блокируется на фоне возвратного торможения. По-видимому, обе группы синапсов находятся в непосредственной близости друг от друга, и синапсы возвратного ТПСП оказывают шунтирующее влияние на синапсы возвратного ВПСП.
- Близкое расположение синапсов этих двух систем доказывает, что в КК, где отсутствует возвратное торможение, значительно облегчена генерация возвратного возбуждения и при увеличении силы ортодромного раздражения в них возникает новая волна ВПСП, по-видимому, также возвратной природы.
Соотношение внутриклеточных реакций нейронов ОЛ лягушки с обонятельными ВВ
Одновременная внутриклеточная и фокальная регистрация ВВ показала, что в большинстве зарегистрированных нейронов на адекватное раздражение возникают колебания мембранного потенциала, синхронные с ВВ. Чаще всего во время развития суммарных ВВ удается зарегистрировать гиперполяризационные колебания мембранного потенциала, реже наряду с гиперполяризационными — деполяризационные. У ряда вторичных нейронов наблюдалось лишь медленное гиперполяризационное отклонение потенциала с большим латентным периодом. Оно совпадало с серединой веретена ВВ и не блокировало потенциал действия. Искусственная гиперполяризация нейронов показала, что у большинства из них амплитуда колебаний уменьшалась, а у других увеличивалась. Все это свидетельствует о том, что в большинстве нейронов во время адекватного раздражения доминируют ТПСП, однако ВПСП также, несомненно, принимают участие в формировании колебаний мембранного потенциала, синхронных с ВВ.
Следует отметить, что между МК и КК наблюдалось отличие, в КК значительно чаще на ВПСП генерировался спайк.
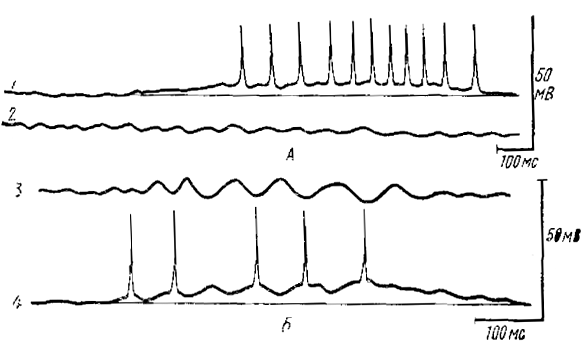
раздражение в ОЛ лягушки (по Гусельниковой и Гусельникову, 1975)
А — внутриклеточная регистрация ГК (1) и суммарные ВВ (2); В — суммарные ВВ (3) (отрицательность вверх), и внутриклеточная регистрация МКК (4) (50 мВ для внутриклеточной записи). Суммарные ВВ на А и Б регистрируются в ростральной части обонятельной луковицы
Длительная гиперполяризация в некоторых ВН могла увеличиваться на фоне искусственной гиперполяризации, следовательно, она не является ТПСП, а, вероятно, отражает блок ВПСП, о котором говорилось выше. В ГК во всех случаях возникала с большим латентным периодом тоническая деполяризация со спайками (рис. 8, А), потенциалы действия были синхронны с ВВ только в начале разряда. В МКК развивался обычно коротколатентный ВПСП со спайками, которые всегда бы ли синхронны с позитивной фазой фокальных ВВ (рис. 8, Б) в отличие от вторичных нейронов, разряд которых всегда коррелировал с негативной фазой. Уже эти данные показывают, что ГК отвечают с большим латентным периодом (до 500 мс), когда веретено достигает своего полного развития и, следовательно, ГК не могут принимать участия в «запуске» веретена. Поскольку ГК являются интернейронами возвратного торможения, то, следовательно, последнее не формирует ВВ, как это имеет место в таламических нейронах при развитии σ-ритма.
МКК, которые являются интернейронами раннего торможения, активно участвуют в формировании ВВ; об этом свидетельствует короткий латентный период ВПСП и потенциалов действия и строгая приуроченность последних к позитивной фазе фокальных ВВ (рис. 8, Б).
Но этих данных пока мало, чтобы сделать заключение о механизмах генерации ВВ. С этой целью мы исследовали механизм ритмического разряда последействия (РРП), который возникает на сильное ортодромное раздражение. Так как синаптические процессы, возникающие на ортодромное раздражение, достаточно подробно изучены, РРП легче подвергнуть анализу.
РРП, возникающий на ортодромное раздражение в ОЛ на фоне позитивной фазы ортодромного вызванного потенциала, есть, по-видимому, не что иное, как спровоцированная электрическим раздражением вспышка ВВ.
Это подтвердилось многочисленными исследованиями нашей лаборатории (Энговатов,1971; Гусельникова, Гусельников, 1975 и др.).
Одновременная регистрация внутриклеточной активности нейронов ОЛ и РРП на ортодромное раздражение и спонтанных ВВ дали возможность сделать следующие общие заключения.
- Основная роль в формировании РРП и ВВ принадлежит ТПСП, так как часто во ВН наблюдается только последовательность ТПСП при генерации этих ритмических процессов и на фоне искусственной гиперполяризации мембраны ВН током, колебания МП уменьшаются.
- По временным характеристикам и общей картине эти ТПСП являются ранними ТПСП (рис. 9), но не возвратными, что подтверждается тем фактом, что разряд МКК (интернейронов раннего ТПСП) коррелирует с положительной фазой волн РРП и ВВ, тогда как разряд ГК (интернейрона возвратного ТПСП) возникает с большим латентным периодом, падает на середину веретена и играть роль в запуске последнего не может. Однако первые разряды ГК совпадают с синхронным разрядом остальных клеток и приходятся на негативную фазу волны, что говорит о некотором вкладе ГК в формирование ВВ, но главная роль их, видимо, другая.
- Во время РРП и ВВ наблюдаются деполяризационные колебания во ВН. Эти колебания не являются результатом постанодальной экзальтации после ТПСП, так как в ряде случаев во время РРП и ВВ могли наблюдаться одни деполяризационные колебания мембранного потенциала без предшествующего ТПСП.
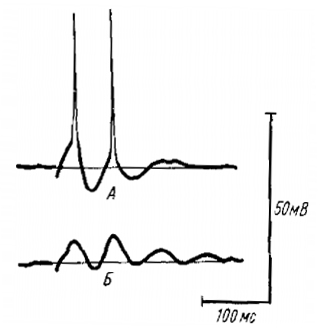
Это деполяризационное колебание не является и афферентным ВПСП, анализ свойств дает основание отождествлять его с возвратным ВПСП. Так, например, гиперполяризация мембраны ВН током увеличивает этот ВПСП (см. рис. 9), так как это характерно для возвратного ВПСП, но не афферентного. Он уменьшается при развитии возвратного ТПСП, в то время как афферентный ВПСП обычно увеличивается.
Таким образом, последовательность явлений, приводящая к генерации ВВ и РРП, нам представляется следующим образом.
Афферентный залп, приходящий по обонятельному нерву, вызывает активацию апикальных дендритов ВН, возникает афферентный ВПСП, который генерирует спайк, распространяющийся к соме ВН.
➥ Более подробно: Физиология дендритов нервных клеток и их функциональная роль
В это же время происходит активация МКК. интернейрона раннего ТПСП посредством дендро-дендритного синапса между дендритами ВН и МКК. В результате активации МКК на апикальных дендритах ВН возникает ранний ТПСП, а спайк сомы вызывает активацию возвратной активирующей системы, оканчивающейся на базальных дендритах, где возникает возвратный ВПСП с латентным периодом, соответствующим концу развития раннего ТПСП.
Возвратная активирующая система значительно более эффективна в КК и, как сообщалось выше, возвратный ВПСП регистрируется во всех КК без исключения; причем на ВПСП часто генерируется ПД в отличие от МК.
Важно напомнить также, что возвратная активирующая система КК мало пересекается с возвратной активирующей системой МК. Следовательно, за счет МК и в большей степени КК происходит (посредством латеральных связей) широкое вовлеченье новых ВН.
Это хорошо согласуется с морфологией, так как известно, что КК значительно больше, чем МК, даже у лягушки (Хмелевская. Гусельникова, 1973). Аксонные коллатерали обоих типов клеток идут по направлению к наружному плексиформному слою, где они контактируют с дендритами как КК, так и МК. Известно также, что дендрита КК и МК образуют синапсы с дендритами МКК. Поэтому, с одной стороны, мы наблюдаем облегчение вероятности появления повторною раннего ТПСП во ВН на фоне возвратного ВПСП за счет того, что последний, по-видимому, электротонически распространяется на апикальные дендриты, повышает их возбудимость и способствует генерации спайка на длящееся тоническое афферентное возбуждение, которое вновь активирует через дендро-дендритные синапсы МКК. С другой стороны, возможно непосредственное возбуждение МКК КК за счет того, что возвратный ВПСП в них часто генерирует спайк, который распространяясь в антидромном направлении может возбуждать МКК также через дендро-дендритные синапсы.
Возбуждение по этим путям новых МКК подразумевает широкое вовлечение в ритмику ранее не охваченных возбуждением ВН. Подтверждением тому, что КК «работают» частично независимо от МК, может служить тот факт, что на повторное ортодромное раздражение при небольших интервалах между раздражениями, когда второй стимул падает на деполяризацию от первого, в КК обычно возникает ранний ТПСП, тогда как в МК на повторное раздражение (когда в МК регистрируется возвратное торможение) ранний ТПСП не регистрируется.
Вероятно, десинхронизирующим механизмом веретена ВВ отчасти является возвратная тормозная система, которая в силу высокого порога включается с большим латентным периодом примерно в середине веретена. Первые несколько спайков ГК возникают синхронно с разрядом других клеток на негативной фазе ВВ, по активность ГК обусловливает развитие возвратного ТПСП на базальных дендритах МК, что приводит к шунтированию синапсов возвратного возбуждения, расположенных здесь же на базальных дендритах. И именно тот факт, что в большинстве нейронов во время адекватного раздражения регистрируется медленное гиперполяризационное отклонение потенциала с большим латентным периодом, которое увеличивается на фоне искусственной гиперполяризации мембраны нейрона, говорит о том, что блок ВПСП является доминирующим фактором, а не возвратный ТПСП в «дезорганизации» системы ранний ТПСП — возвратный ВПСП, это в конечном счете и приводит к прекращению ритмики. Из наших данных следует, что сами по себе КК не в состоянии генерировать ритмический устойчивый процесс. Поэтому, когда в МК возникает возвратное торможение и блок возвратного возбуждения, в КК наступает дефицит возбуждения, уровень которого ниже необходимого для поддержания ритмической активности и в конечном итоге имеет место «разрушение» веретена ВВ.
Таким образом, механизм формирования ритмической активности в ОЛ отличается от механизма, постулируемого гипотезой Андерсена и Экклса для объяснения σ-ритма в таламусе млекопитающих, где основная роль принадлежит тормозному фазированию таламических нейронов, осуществляемому системой возвратного торможения.
Сходство между механизмами формирования веретенообразной ритмической активности таламуса млекопитающих, согласно Андерсену и Экклсу и предложенному нами для ВВ в ОЛ лягушки, заключается в том, что фазирование осуществляется тормозной системой, но в нашем случае эта система раннего торможения (хотя по отношению апикальных дендритов ВН это торможение можно рассматривать как возвратное).
Второй особенностью нашей гипотезы является предложенный механизм повышения возбудимости во ВН после раннего ТПСП, который осуществляется системой возвратного возбуждения, хорошо развитой у КК. В теории Андерсена и Экклса этот вопрос остался неясным.
И, наконец, третья особенность заключается в том, что, по нашему мнению, дезорганизация ритмического процесса осуществляется активным включением системы возвратного торможения и возникновением блока возбуждения во ВН, наряду с остаточным характером афферентного возбуждения, тогда как в таламических нейронах роль дезорганизующего фактора отводится, видимо, пассивному накоплению остаточной гиперполяризации.
Возникает вопрос, приложимы ли наши данные, полученные на примере анализа ВВ в ОЛ лягушки, к механизмам такой же активности в ОЛ у других животных а в первую очередь рептилий и млекопитающих.
Наши исследования на ОЛ рептилий (ящерица, черепаха) совпали с данными, полученными на лягушке. Косвенные данные на немлекопитающих получил Грейстоун и сотрудники (Graustone et аl., 1970), которые показали, что частота ВВ в ОЛ жаб, игуан и змей зависит от температуры сходным образом, что они связывают с общим генезом ВВ у этих животных. Что касается млекопитающих, то вопрос о генезе ВВ в ОЛ пока остается открытым. Однако последние данные, полученные нами на ОЛ крысы, заставляют полагать, что и здесь, возможно, имеет место сходный .механизм генерации ВВ.
Необходимо также отметить, что ОЛ является пейсмекером обонятельной ритмической активности в высших обонятельных центрах. Выше мы указывали, что почти у всех представителей различных классов позвоночных под влиянием обонятельной импульсации в центральных структурах обонятельного анализатора возникают обонятельные волны. Идентичны ли они по своему происхождению, ВВ и ОЛ сказать трудно.
Будро (Boudreau, 1964) с помощью цифровой вычислительной машины провел специальный анализ биоэлектрической активности ОЛ и пириформной коры кошки при их одновременной регистрации. Он пришел к выводу, что обонятельные волны пириформной коры возникают в результате поступления в кору импульсов по латеральному обонятельному тракту от ОЛ, причем эти импульсы следуют в ритме волн самой обонятельной луковицы. Однако данные Будро еще не доказывают, что генерация волн в пириформной коре не может осуществляться самостоятельно. Эйдельберг (Eidelberg, 1963), например, считает, что ВВ генерируются в ОЛ, пириформной коре и миндалине кошки независимо друг от друга. Наши данные (Гусельников, 1964) подтвердили наличие раздельных механизмов генерации ВВ в гиппокампальной коре и обонятельной доле черепахи, однако последняя, видимо, может являться водителем для коркового ритма.
Таким образом, нейрофизиологический механизм генерации ВВ в других обонятельных структурах мозга изучен недостаточно, для его выяснения требуются дальнейшие исследования.