Общая классификация электрической активности головного мозга является сложной задачей, в то же время совершенно очевидно, что необходимость такой классификации давно назрела, так как в общей электрофизиологии мозга и клинической электроэнцефалографии списан широкий спектр различных ЭЭГ феноменов и определенное их упорядочивание и классификация необходимы. Здесь возникает ряд трудностей и прежде всего правомочность классификации, казалось бы, сходных ЭЭГ феноменов, описанных у животных и человека, т. к. мы еще не всегда имеем право относить их к одному классу явлений. Другая трудность заключается в том, что на ряд биоэлектрических процессов мозга в литературе часто существуют диаметрально противоположные взгляды, совместить которые не всегда возможно. И, наконец, наибольшие сомнения возникают там, где совершенно не ясен генез тех или иных электрических явлений, а известны только общие внешние характеристики. Поэтому и разбираемая здесь классификация, не будет лишена недостатков.
Предложенная классификация является расширенным и измененным вариантом уже имеющихся классификаций (Соколов, 1962; Кожевников, Мещерский, 1963). Мы остановимся на всех видах суммарной электрической активности мозга.
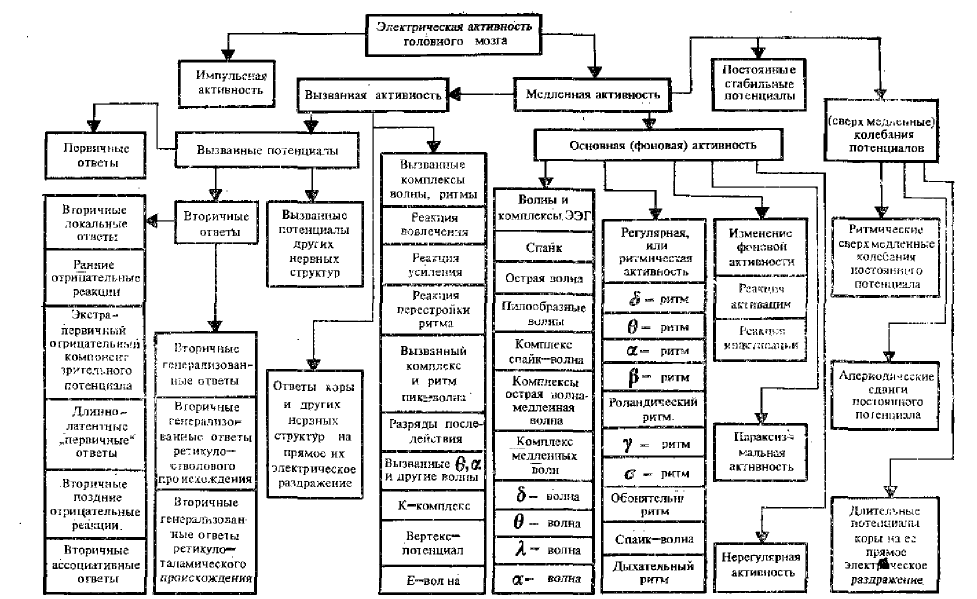
Всю биоэлектрическую активность головного мозга можно разделить на две крайние группы: импульсную активность и суммарную медленную активность (см. схему общей классификации различных видов электрической активности головного мозга).
Импульсная активность представляет собой в основном форму деятельности аксонов, тел нервных клеток и частично дендритов.
Медленная активность определяется, как мы уже говорили ранее, в основном алгебраической суммой ВПСП и ТПСП сомы и дендритов нервных клеток, а также, видимо, сдвигами метаболических процессов мозга и активностью глиальных элементов.
Медленная суммарная активность головного мозга делится на три основные, группы:
- Вызванную электрическую активность мозга;
- Основную («спонтанную», фоновую, автономную) активность;
- Медленные и сверхмедленные колебания потенциала.
К вызванной электрической активности головного мозга относятся простые и сложные (комплексные) колебания биопотенциалов, возникающие в коре и различных других образованиях головного мозга при прямом электрическом, одиночном или ритмическом их раздражении и в ответ на поступление в них импульсов по различным афферентным путям при адекватной или электрической стимуляции. Они объедины под общей рубрикой — вызванные потенциалы.
К вызванным потенциалам относятся:
- Первичные ответы коры больших полушарии;
- Вторичные ответы коры больших полушарий;
- Вызванные потенциалы других нервных центров (например, гиппокампа, мозжечка, таламических структур и т. д.);
- Ответы коры и других нервных структур на прямое их электрическое раздражение.
Кроме того, к вызванной активности отнесены спровоцированные различными способами комплексы, волны и ритмические электрические процессы, которые в норме характерны для спонтанной суммарной активности того или иного отдела мозга.
К вызванным комплексным реакциям, волнам и ритмам относятся:
- Реакция вовлечения;
- Реакция усиления;
- Реакция перестройки ритма;
- Вызванный комплекс и ритм пик-волна;
- Разряды последействия;
- Вызванные θ-, α-, обонятельные волны и т. д;
- К-комплекс, вертекс-потенциал, Е-волна (волна «внимания»).
Дадим краткую характеристику всем видам активности.
Первичные ответы коры больших полушарий. К ним относятся ответы, возникающие только в корковых проекционных зонах анализаторов животных и человека при сенсорных адекватных раздражениях или при электрическом раздражении рецептора, проводящих специфических путей и релейных ядер.
Вторичные ответы коры больших полушарий возникают при адекватных и неадекватных раздражениях рецептора, а также определенных групп таламических ядер и ретикулярной формации ствола мозга.
Электрогенез вторичных ответов отличается от такового первичных ответов. Вторичные ответы можно разделить на:
- Вторичные локальные ответы, куда относятся: а) ранние отрицательные реакции, б) экстрапервичный отрицательный компонент зрительного потенциала, в) длиннолатентные «первичные» ответы, г) вторичные поздние отрицательные реакции, д) вторичные ассоциативные ответы, е) вторичные коллазальные ответы;
- Генерализованные ответы, куда относятся: а) вторичные генерализованные ответы ретикуло-стволового происхождения и б) вторичные генерализованные реакции ретикуло-таламического происхождения.
Вызванные потенциалы других нервных центров
Электрогенез этих видов реакций (так же как и в коре больших полушарий) целиком определяется характером афферентных связей и строением центров. Они возникают также на адекватное или электрическое раздражение рецепторов, проводящих путей или центров, с которыми исследуемый центр имеет связи. Так, например, вызванные потенциалы мозжечка возникают на световую, звуковую, сомато-сенсорную стимуляцию, на прямое электрическое раздражение тех же рецепторов, ряда таламических ядер и других подкорковых ядер, а также на прямое электрическое раздражение отдельных зон коры больших полушарий.
Реакция вовлечения (recruiting response) состоит из серии медленных отрицательных волн, возникающих с большим латентным периодом в коре больших полушарий, при ритмическом, низкочастотном (5—10 в 1 с) электрическом раздражении ядер медиального таламуса.
Впервые описана и исследована Демпси и Морисоном (Dempsey, Morison, 1942).
➥ Основная статья: Реакция вовлечения и отвлечения
Реакция усиления (augmenting response) возникает в коре больших полушарий при низкочастотном раздражении специфических ядер таламуса и волокон радиации. Описана в 1943 г. теми же исследователями.
Реакция перестройки ритма впервые была описана. Эдрианом и Мэттьюзом (Adrian, Mattews, 1934) в электроэнцефалограмме человека. Они называли ее «реакцией мельканий» (flicker response). В дальнейшем эту реакцию разные авторы стали называть «реакцией навязывания ритма, следования ритма и усвоения ритма». Мы будем пользоваться термином «реакция перестройки ритма».
Эта реакция заключается в изменении, перестройке спонтанной активности коры мозга под влиянием, как правило, световых ритмических раздражений сетчатки глаза. В этом случае электрическая активность коры приобретает ритмический характер с частотой, равной или в целое число раз большей или меньшей по сравнению с частотой применяемого ритмического светового раздражения.
Реакция перестройки ритма наблюдается и при ритмических афферентных раздражениях других модальностей. Например, описана реакция перестройки в ЭЭГ человека при действии ритмического звука (Davis et al., 1939; Xeher, 1961).
В ЭЭГ животных впервые реакция перестройки ритма на ритмический световой раздражитель описана и исследована Бартли (Bartly, 1936) и М. Н. Ливановым (1940).
Комплекс спайк-волна и ритм спайк-волна
➥ Основаня статья: Спайк-волна ЭЭГ
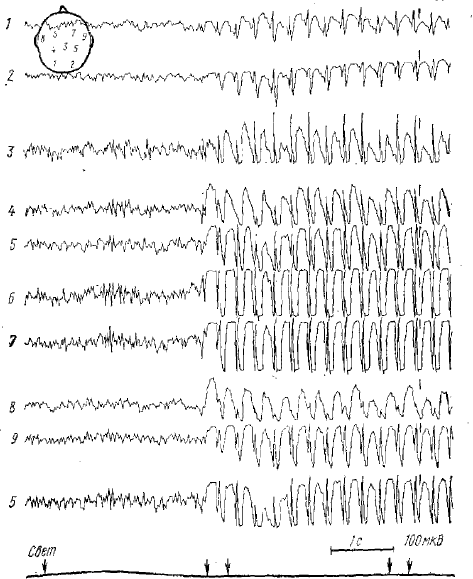
1—9 — точки отведения ЭЭГ на cxeме сверху
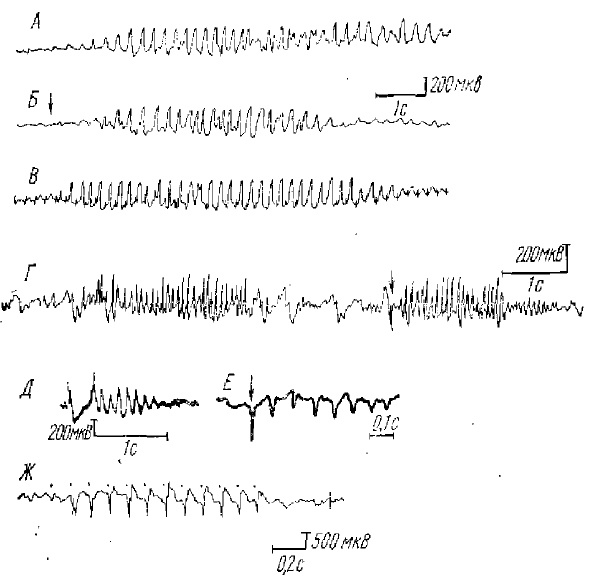
Комплекс спайк-волна является патологическим комплексом в ЭЭГ животных и человека и характерен для специфического случая генерализованной эпилепсии, носящей название «petit mal», или «малого миоклонического припадка» (рис. 1). Характеризуется комбинацией спайка и следующей за ним медленной волны длительностью от 200 до 500 мс. В литературе встречается и другой термин для этого комплекса — спайк-купол (spike-deme). Как правило, во время клинического припадка petit mal эпилепсии в ЭЭГ возникает ритм спайк-волна, который состоит из ритмически повторяющихся с частотой 2,5—3,5 кол/с патологических комплексов спайк-волна. Поскольку наиболее выраженным компонентом комплекса при petit mal —эпилепсии является волна, а не пик, то некоторые исследователи (Кругликов и др., 1970) предлагают назвать комплекс не спайк-волна, а волна-пик.
Комплекс спайк-волна и спайк-волна ритм могут быть вызваны искусственно у животных рядом способов, путем стимуляции неспецифических таламических ядер химически (введением алюминиевой пасты) или с помощью электрического раздражения тех же ядер с частотой 3 в 1 с.
Разряды последействия
Возникают в ЭЭГ при различных одиночных адекватных раздражениях того или иного анализатора путем прямой электрической стимуляции ряда ядер таламуса или коры. Разряды последействия описаны и изучены у животных (Bartly. Bishop, 1933; Adrian, 1941; Bremer, 1949; Chang, 1951; Брейзье,. 1952; Гусельников. Супин, 1962, 1968, и др.) и у человека (Cigancc, 1958; Barlou, 1960; Cohn, 1964, и др.)
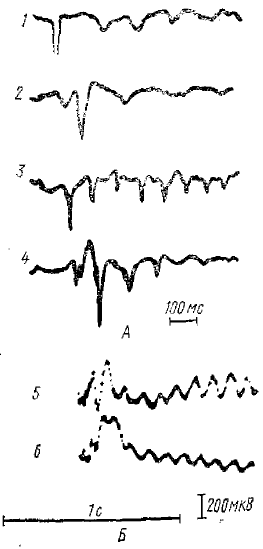
1 — ответ передней эктосильвиевой извилины на звуковой щелчок, 2— ответ латеральном извилины на свет, 3 и 4 — то же, что на 1 и 2 при другом наркозе, 5 и 6—РРП получены на одиночную световую вспышку путем автоматического усреднения 600 предъявлений раздражителя, (5 — отведение от правого полушария, 6— от левого). Момент светового раздражения соответствует началу записей
Все разряды последействия имеют, как правило, вид вспышки ритмических электрических колебаний в коре (или таламическом ядре) и возникают после начального ответа на применяемое раздражение, например, после первичного ответа в коре в случае афферентного раздражения или дендритного ответа в случае прямого Электрического раздражения коры и т. д. (рис. 2). Обычно вспышка волн ритмического разряда последействия (РРП) носит затухающий характер; вначале амплитуда волны максимальна, к концу постепенно убывает. Частота волн PPП различна у различных животных и человека. У человека она близка к частоте α-ритма (10— 12 Гц), у кошек около 10 Гц (в барбитуровом наркозе). По Эдриану (Adrian, 1941), частота РРП у лягушки может достигать 20 Гц, у кролика (Bartley, Bishop, 1933) около 5 Гц и т. д.
Разрядами последействия ряд исследователей называют и другие ЭЭГ явления — вспышки высокой частоты неизвестной природы. Так, например, Бремер и Бонне (Bremer, Bonnet, 1950) описали так называемый «быстрый разряд последействия» с частотой до 60 Гц, а Эдриан, Джаспер, Айджмон-Марсан и сотрудники (Adrian, 1941; Ajmone-Marsan, Stoll, 1951; Jasper et al., 1952) списали «судорожный разряд последействия», возникающий в структурах мозга после их длительной и сильной стимуляции.
В ряде случаев отмечается, что РРП имеют локализованный характер: возникают в специфическом релейном таламическом ядре и соответствующей ему проекционной зоне коры мозга. При одиночном раздражении неспецифических ядер таламуса также возникают РРП, более широко распространенные по коре.
Вызванные волны
Вызванные волны (ВВ)— явление, видимо, очень близкое, а в ряде случаев, как мы убедимся далее, часто тождественное разряду последействия. Условия их возникновения и локализация, как правило, совпадают с таковыми для РРП. Под ВВ подразумевается процесс провокации в электрокортикограмме (ЭКоГ) тех или иных характерных волн, которые в норме могут характеризовать основной рисунок спонтанной активности мозга. Так, например, во время стадии начальной фазы сна у человека и животных внешнее афферентное раздражение (вспышка света) может спровоцировать появление в ЭКоГ α-волны, являющейся компонентом α-ритма, характерного для ЭЭГ спокойного состояния человека. Та же вспышка может вызвать и целый цуг α-волн (вспышка α-активности), в этом случае говорят о появлении РРП. То же относится и к некоторым другим видам ВВ.
К-комплекс и вертекс-потенциал (Vertex, V-potential)
➥ Читайте также: Неэпелиптеформная активность на ЭЭГ
Видимо, правильнее было бы рассматривать вместе с вторичными ответами коры больших полушарий. Но так как эти ответы описаны под этим названием лишь для ЭЭГ человека, а механизмы их спорны, имеет смысл рассмотреть их отдельно. Кроме того, К-комплекс может возникать и спонтанно.
Вертикальные линии — отметка раздражения
К-комплекс состоит из двух-трехфазной острой волны с последующим разрядом последействия (σ-послеразряд) (рис. 3). Вначале полагали, что этот ответ является специфическим ответом коры, так как он возникал в ЭЭГ в начальной стадии сна у человека только на звуковое раздражение и регистрировался локально в теменной области полушарий.
Однако дальнейшие работы (Кац, 1959; Пучинская, 1963, 1967, и др.) показали, что К-комплекс возникает при самых разнообразных раздражениях и может наблюдаться не только в начальной стадии сна. Он был отнесен к неспецифическим вызванным потенциалам. К-комплекс возникает в определенной стадии сна и спонтанно, нередко сопровождается вегетативными и двигательными реакциями и не наблюдается во время парадоксального сна (Шаповальников, 1971).
Ряд исследователей придают К-комплексу важное функциональное значение, рассматривая его как электрографическое отражение активности специального антипробуждающего механизма. Есть даже попытка связать К-комплекс со зрительными образами во сне: чем больше проявляется К-комплексов на звук у человека во сне, тем больше и ярче зрительные образы во сне (Fernander, 1969). Некоторые физиологи электрогенез К-комплекса связывают с механизмами, лежащими в основе petit mal эпилепсии.
Вертекс-потенциал впервые описал Девис (Davis, 1939; Devis et al., 1938), затем Гасто (Gastaut, 1951, 1952), Банко (Bancaud et al., 1953) и др. Он регистрируется в области макушки (вертекса) черепа (т, е. там же, где и К-комплекс). При действии различных раздражителей обычно он выражен либо в виде острого пика или медленной однофазной или двухфазной волны (см. рис. 3). Такой нестабильный характер формы ответа (Кац, 1958, 1959; Русинов, 1969, и др.) зависит, видимо, от повторности применяемого раздражения. Так, В. С. Русинов (1969) указывает, что в процессе применения раздражителя (например, звука), прежде чем вертекс-потенциал приобретает форму пика, он проходит стадию низкоамплитудной медленной волны. При дальнейших применениях раздражителя он «растягивается» во времени. Изучая динамику изменения зертекс-потенциала по мере применения раздражителя, отмечая также широкие вариации его формы у разных испытуемых и даже у одного и того же испытуемого во время одного сеанса исследования, Кац приходит к выводу, что и К-комплекс является одной из вариаций этого ответа. С этих позиций К-комплекс и вертекс-потенциал являются вариациями одного и того же ответа — неспецифического ответа коры больших полушарий человека.
Вертекс-потенциал был сравнен со вторичным генерализованным по коре неспецифическим ответом Форбса и Морисона. Затем это мнение было отвергнуто на том основании, что вертекс-потенциал локален. Однако в работах В. С. Русинова с сотрудниками и других исследователей было показано, что вертекс-потенциал является действительно неспецифичным ответом, он возникает на самые различные раздражения, имеет большой латентный период (70—90 мс) и регистрируется не только в области вертекса, но и в других областях коры.
Так, Кац (1959) наблюдал явление распространения ответа на другие области коры больших полушарий, что сопровождалось нарастанием его амплитуды. Данные лаборатории В. С. Русинова, а также данные Ларссона (Larsson, 1960) опровергают высказанную ранее точку зрения Освальда (Oswald, 1959), что неспецифический ответ является артефактом и возникает в результате сдвига кожи на черепе при движении ушей. Неспецифический ответ коры больших полушарий человека может быть использован как ЭЭГ показатель неспецифических влияний на кору больших полушарий. Примером использования, этого ответа служат работы лаборатории В. С. Русинова (Кац. 1959; Пучинская, 1963, 1967; Соколова, 1954; Русинов, 1969, и др.). Что касается ЭЭГ животных, то о неспецифическом ответе, видимо, гомологе неспецифического ответа коры человека, мы будем говорить в специальном разделе нашего курса. Укажем только, что К-комплекс (или вертекс-потенциал) описан и у обезьян (Cohn, 1969; Bert et al., 1969).
Е-волна
Волна «ожидания», или Е-волна, описана Уолтером (Walter, 1963) и названа им «когерентной негативной вариацией» (КНВ). Возможно, она является измененным вертекс-потенциалом в процессе выработки временной связи. Генез ее остается пока неясным. Топография Е-волны ограничена передней и центральной областями головы, включая Фронтальную, моторную и соматосенсорную зону. Е-волна следует сразу за условным сигналом, развивается очень медленно и имеет большую длительность.

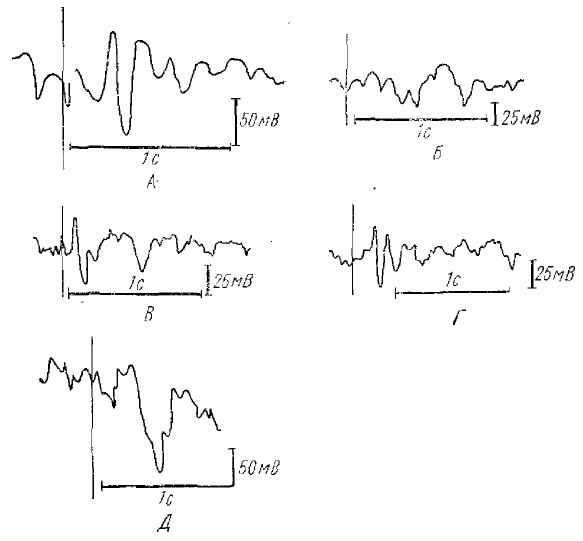
А — изолированно примененный щелчок; Б — изолированно примененные вспышки; В — применение щелчка вместе со вспышками; Г—волна на условный сигнал — щелчок (по Уолтеру, 1965)
Уолтером (1965) она была найдена в следующем опыте. Применялись два раздражителя — звуковой щелчок и ритмический свет — с интервалом в 1 с (рис. 4). Изолированно примененный щелчок вызывает положительную волну (рис. 4, А), замаскированную большим отрицательным компонентом. Ответы на изолированные серии вспышек содержат несколько кратких отрицательных компонентов, также наложенных на небольшое положительное колебание (рис. 4, Б). Когда щелчки применяются вместе со вспышками с интервалом в 1 с (рис. 4, В).
Каждая запись—усредненные ответы на 12 предъявлений раздражителя. Отклонение вверх соответствует электронегативности оба ответа воспроизводятся с небольшими изменениями. После этого испытуемый получает инструкцию нажимать на кнопку в ответ на второй раздражитель (во время вспышек света), что прекращает вспышки, которые являются безусловным подкреплением. В этом случае, в ответ на условный сигнал — щелчок— возникает большая отрицательная волна, которая и была названа Е-волной, или волной «ожидания» (испытуемый ожидает вспышек света, чтобы их включить, нажав на кнопку). Такая картина наблюдается длительное время (рис. 4, Г), пока внимание испытуемого сохраняется, и он точно нажимает на кнопку. Однако, если исключить подкрепление (вспышки света), Е-волна полностью исчезает.
Е-волна возникает сходным образом и в экспериментах, в которых испытуемый должен принять решение в уме, вместо того чтобы выполнять двигательный акт. Необходимо только, чтобы он проявил некоторое умственное усилие в ответ на второй раздражитель (мысленно представил нажатие на кнопку).
Уолтер полагает, что Е-волна имеет корковую природу, так как она инвертирует, если электрод, введенный в мозг человека, проходит через поперечник коры. Это свидетельствует, по его мнению, об ее происхождении в поверхностном плексиформном слое коры. Он также считает, что она отражает деполяризацию сети апикальных дендритов в результате разрядов пирамидных нейронов и не исключает роли гуморальных факторов в ее происхождении.
Некоторые исследователи полагают, что волна «ожидания» имеет место и в ЭЭГ у животных. Так, например, Камп и Ван Риян (Kamp, Rijan, 1970) показали, что при выработке у собаки пищевого двигательного условного рефлекса похожий негативный сдвиг потенциала имеет место в ЭЭГ в период, предшествующий получению пищи.
Основная (фоновая, спонтанная) электрическая активность головного мозга
По внешнему характеру основную активность мозга можно разделить на три вида (см схему):
- Регулярную, или ритмическую, активность, характеризующуюся длинной серией тех или иных волн с незначительной вариацией их частоты;
- Нерегулярную, или аритмическую, активность, состоящую из волн различной длительности л амплитуды;
- Пароксизмальную активность, возникающую на фоне обычной спонтанной активности в виде определенных групп высокоамплитудных волн и комплексов, амплитуда которых внезапно нарастает и спадает.
Однако, прежде чем рассмотреть все эти виды активности, дадим общую характеристику волнам и комплексам, которые либо могут целиком составлять основную активность, либо периодически появляться на ее фоне.
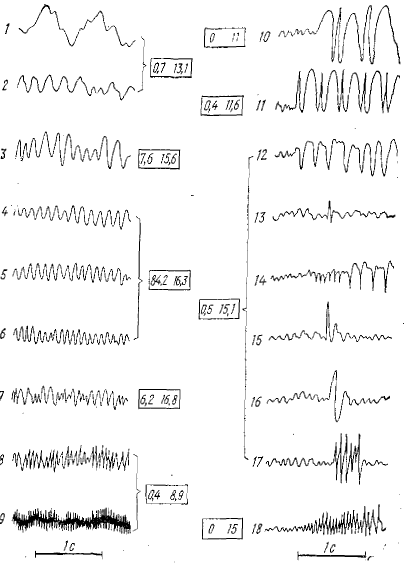
1 — очень медленная активность 0,5—5 кол/с, 2 — медленная активность 3— 6 кол/с, 3 — незначительно замедленная активность 6—8 кол/с, 4, 5, 6 — нормальная активность от 8,5 до 12 кол/с, 7 —учащенная активность. 8 —быстрая активность, 9 — очень быстрая активность, 10 —pelit mal (частота 2 цикл.) в 1 с), 11— petit mal (частота 3 цикла в 1 с), 12 — психомоторные судороги, 13 — низкоамплитудные острые спайки, 14 - положительные спайки с частотой, 14 и 6 кол/с, 15 — высокоамплитудные спайки, 16 — высокоамплитудные замедленные спайки, 17 — множественные спайки; 18 — grand mal — эпилепсия (12—13 кол/с). Цифры в квадратиках % времени, которое тот или иной вид активности может занимать в ЭЭГ здорового (слева) и больного (справа) человека
Острая волна. Впервые была описана Джаспером (Jasper, 1941). Спорадические быстрые колебания, с быстрой крутизной нарастания и более пологой крутизной нисходящей части. Длительность волны лежит в пределах 80—200 мс (рис. 5).
Спайк. Спорадические быстрые колебания длительностью около 20 мс с быстрой крутизной нарастания и спада восходящей и нисходящей фазы. При отведении с поверхности мозга длительность спайков может равняться 10 мс. Спайки с длительностью 5—10 мс И. С. Беритов и сотрудники (1943) обозначили как иглоподобные волны. Как острые волны, так и спайки могут возникать внезапно, через нерегулярные интервалы на фоне основной активности. Они могут быть положительные или отрицательные, одиночные и множественные, двухфазные и монофазные, нерегулярные и ритмические (см. рис. 5).
Группы спайков и острых волн определяются как разряды или вспышки. Как правило, спайки и острые волны связаны с патологией, хотя могут иметь место и в обычной ЭЭГ животного и человека.
Комплекс спайк-волна. О нем мы уже говорили ранее. Необходимо только отметить, что могут иметь место вариации этого комплекса, например, две спайк-волны или множественные спайк-волны, острая и медленная волна, когда спайк достигает длительности 80—200 мс, а медленная волна 500—1000 мс.
Дельта (δ)-волны. Длительность δ-волн составляет 250—500 мс. Если они собраны в регулярные серии, то составляют ритм электрической активности мозга — δ-ритм (см. рис. 7).
Комплекс медленных волн. Группа из двух и более медленных волн, повторяющаяся с определенным постоянством формы (см. рис. 5).
Пилообразные волны. Повторяющиеся ритмические острые волны определяют как пилообразную активность, или пилообразные волны (см. рис. 5).
Всем перечисленным выше видам волн и комплексам большое внимание уделяется в клинической ЭЭГ. Спайки, острые и пилообразные волны и комплексы типа спайк-волна объединяют под названием судорожных, или эпилептиформных, потенциалов. На рис. 5 дана классификация электрической активности ЭЭГ человека (Gibbs, 1950). Левая колонка (1—9) показывает различные типы ритмической активности мозга человека, а правая — эпилептическую активность. В пределах цифр от 3 до 7 нормальная ЭЭГ человека и слабые отклонения от нормы (3 и 7). На 1 и 9 крайние степени, на 2 и 8 — значительная степень патологии. ЭЭГ здорового человека почти лишена эпилептических потенциалов (допускается содержание небольшого количества медленных и быстрых волн). Цифры, ограниченные квадратами левого столбца, показывают процент времени, которое тот или иной вид активности может занимать в ЭЭГ взрослого здорового человека; у правого столбца — процент времени, которое занимает данная активность в ЭЭГ больных эпилепсией.
Предложены и другие способы классификации ЭЭГ здорового и больного человека.1
Характеристика волн, встречающиеся в виде одиночных волн или составляющие ритмы электрической активности нормальной ЭЭГ животных и человека
Тэта-(θ) волны. Их длительность составляет 150— 200 мс. Имеют место как в ЭЭГ человека, так и животных, хотя происхождение их, видимо, различно.
Ламбда-(λ) волны. Одиночные монофазные положительные волны впервые описаны у человека в затылочных областях коры, при том у небольшого процента испытуемых, Гасто и Эвансом (Gastaut, 1951; Evans, 1953). Авторы полагали, что они являются артефактом в результате движения глаз, хотя Грин (Green, 1957) отрицает это и связывает их с механизмом зрения. Длительность λ-волн порядка 300 мс.
Видны λ-волны с амплитудой 100 мкВ, когда собака гуляет вне здания при ярком солнечном свете
Ламбда-волны были описаны в затылочной области коры кошек Родесом и др. (Rodes et al., 1962) и Сторм ван Лёвеном (1965) у собак (рис. 6). Эти исследователи считают, что λ-волны имеют отношение к движению глаз. Предполагается, что они могут служить показателем степени «интереса», с которым животное смотрит. Чем больше количество λ-волн, тем сильнее «интерес».
Альфа-(α) и бета-(β) волны. Длительность α-волн у человека составляет 90—100 мс. Они могут встречаться как в виде отдельных волн, так и составлять регулярный α-ритм. У животных (млекопитающих) в затылочной области коры регистрируются как отдельные альфаподобные полны, так и альфа-подобный ритм (Гусельников, Супин, 1968). Этот вопрос будет рассматриваться нами специально. Что касается β-волн, то они составляют только β-ритм в ЭЭГ человека и у животных не описаны.
Регулярная, ритмическая электрическая активность головного мозга животных и человека (или ритмы электрической активности)
К специфическим ритмам мозга, возникающим спонтанно или при определенных условиях относятся (см. схему): δ-ритм, θ-ритм, α-ритм, β-ритм, роландический, или μ-ритм, σ-ритм (или ритм веретен, веретенообразный ритм), обонятельный ритм, спайк-волна ритм и дыхательный ритм.
Из всех этих ритмов относительно хорошо изучены: α-ритм, θ-ритм, σ-ритм, обонятельный ритм, спайк-волна ритм; элекрогенез других ритмов ЭЭГ изучен слабо.
Ритмы электрической активности складываются из волн, следующих друг за другом с определенным постоянством и имеющих приблизительно одинаковую длительность и форму. Частоты всех ритмов укладываются в диапазон от 1 до 100 кол/с, а амплитуда от 5 до 100— 150 мкВ (последнее относится только к ЭЭГ, регистрируемой с поверхности скальпа или черепа). Ритм, наиболее выраженный в ЭЭГ, в данный момент называют доминантным ритмом.
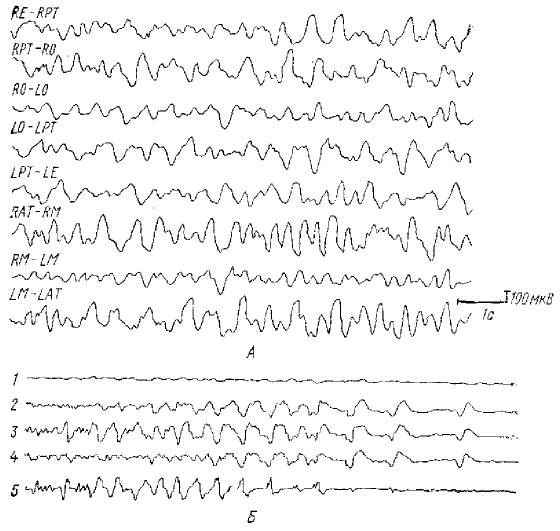
Обозначения отведений ЭЭГ слева М): R — правое полушарие, L — левое полушарие, Е — ушной электрод, РТ—задневисочная область, О — затылочная область, АТ — передневисочная область, М—двигательная область, Р—теменная область, Т — височная область; Б: 1— контрольное отведение между костью лобной пазухи и ухом. Мопонолярное отведение, 2 —- от соматосенсорной коры, 3 — от границы теменной корн и подкоркового белого вещества. 4 — от поверхности теменной коры в той же точке, 5 — транскортикальное отведение (между пунктами отведения 2 и 3); пример ПДВ (запись в пункте 2), СМК — (в пункте 5)
Дельта-ритм. Впервые дельта (δ)-ритм в ЭЭГ человека был классифицирован Уолтером (Walter, 1936), а впоследствии он был описан и у животных, хотя аналогичность их механизмов еще не ясна. Он состоит из ритмических медленных волн длительностью от 250 до 500 мс или даже до 1000 мс. Отсюда, частота его колеблется от 1 до 4 в 1 с (рис. 7). Некоторые авторы полагают, что δ-ритм— ритмическая активность и с частотой 6—8 кол/с называют ее промежуточной медленной активностью. (табл. 1)
δ-ритм наблюдается в ЭЭГ человека, находящегося под наркозом или при поражении кортикальных отделов мозга (опухоль или острый травматический очаг). Максимальная амплитуда его регистрируется тогда на границе опухоли или очага. Это место и носит название δ-фокус. δ-ритм может встречаться и в ЭЭГ здорового человека как во время сна, так и при бодрствовании. Однако в последнем случае амплитуда его не должна превышать 20—30 мкВ (Жирмунская, 1959). Что касается электрогенеза δ-ритма, то пока точных данных на этот счет нет.
Гутман и Милюкас (1969), Гринявичус и соавторы (1969) рассматривают δ-волны как результат изменения поляризации пирамидных клеточных тел коры. Ройтбак (1969) высказывает предположение об аналогии δ-волн с длительными отрицательными потенциалами, возникающими при сильном прямом раздражении коры и допускает участие нейроглии в их генезе.
Долгое время считалось, что δ-активность, возникающая при самых различных физиологических и патологических состояниях, совершенно одинакова. Однако постепенно появились факты зависимости характера этой активности от порождающих ее условий (Gastaut et al., 1961, и др.).
В пределах δ-активности различают более частые и более медленные компоненты. В ЭЭГ выделяют полиморфные и моноритмичные δ-волны, однако в основу существующих классификаций кладут обычно признак распространенности δ-активности по мозгу; в этом случае она разделяется на локальную и генерализованную (Walter, 1936, 1962; Жирмунская, 1963, и др.). До сих пор, однако, не ясны механизмы широкой генерализации этого ритма, определяющие его возникновение и функциональное значение.
На основании экспериментального изучения δ-активности А. М. Гурвич (1969) предлагает разделить ее на известные уже полифазные δ-волны (ПДВ) с частотой 1—3,5 кол/с, обычно непостоянной формы и комплекс стандартной формы, состоящий из негативных волн длительностью 300—500 мс. Последние волны, где бы они ни возникали, отличаются стандартностью формы и были названы стандартным медленным комплексом (СМК) (рис. 7).
Таблица 1. Ритмы ЭЭГ и частотные характеристики по классификации различных авторов (из Кожевникова и Мещерского, 1963)
| Ритм | Частота, кол/с, по Девис (1938) | Частота, кол/с, по Юнгу (1939) | Частота, кол/с, по Джасперу (1941) | Частота, кол/с, по Гибсу и Гибс (1950) | Частота, кол/с, по Хиллу и Пару (1952) | Частота, кол/с, по Шоубу (1951) | Частота, кол/с, по Страуссу, Остоу и Гринштейну (1952) | Частота, кол/с, по П. И. Гуляеву (1960) | Частота, кол/с, по рекомендации Междунарожной федерации обществ электроэнцефалографии и клинической нейрофзиологии |
|---|---|---|---|---|---|---|---|---|---|
| Дельта (δ) | 0,2—4 | 1—7 | 1—6 | 0,5—5 | 0,5—3,5 | 0,5—3,5 | До 6 | 0,5—3 | Менее 4 |
| Тэта (θ) | — | — | — | — | 4—7 | 4—7 | — | 4—7 | 4—8 |
| Промежуточная медленная активность | 4—8 | — | — | — | — | — | 6—8 | — | — |
| Альфа (α) | 8—13,5 | 8—13 | 8—12 | 6—13 | 8—13 | 8—13 | 8—13 | 8—13 | 8—13 |
| Промежуточная быстрая активность | 13,5—20 | — | — | — | — | 14—17 | 13—15 | — | — |
| Бета (β) | Выше 20 | 14—30 | 18—32 | 14—50 | 14—25 | 18—30 | 15—30 | 14—35 | Выше 13 |
| Очень быстрая активность | — | — | — | — | — | Выше 30 | — | — | — |
| Гамма (γ) | Выше 30 | — | 35—55 | — | Выше 26 | — | — | 35—55 | — |
Выраженный θ-ритм наблюдается на 3 и 4 канале в течение первых секунд регистрации. На остальных каналах видны нерегулярные генерализованные θ-волны. Обозначения отведений ЭЭГ слева см. на рис. 7
Тэта(θ)-ритм — складывается из ритмических медленных волн с частотой 4—7 кол/с. Впервые описан и классифицирован как θ-ритм Уолтером и Довейем (Walter, Dovey, 1944). Для ЭЭГ человека принят международный стандарт 4—8 кол/с (рис. 8).
Ряд исследователей полагает, что у человека самостоятельный θ-ритм не существует, и его относят к промежуточной активности между δ- и α-ритмом.
У животных (млекопитающих) θ-ритмом обозначают ритмическую активность с частотой 4—7 кол/с, которая возникает в новой коре, но лучше выражена в лимбических структурах мозга, межуточном мозге и стволе при общей активизации животного, вызванной любыми раздражениями — болевыми, афферентными (при ориентировочной реакции), электрическим раздражением активирующей ретикулярной системы ствола мозга, введением животному антихолинэстеразных препаратов и т. д. Впервые θ-ритм был найден в гиппокамповой коре, где он лучше всего выражен (Gerard el al., 1936; Jung, Kornmiiller, 1938; Green, Arduini, 1953).
Термин θ-ритм (для животных) имеет ряд синонимов: гиппокампальный ритм; гиппокампальный θ-ритм, стресс-ритм и ритм напряжения (Полетаева, Гусельников, 1967). Чаще применяют термин θ-ритм, помня, однако, о том, что аналогичный по частоте ритм ЭЭГ человека мы не можем пока аналогизировать с θ-ритмом животных. θ-ритм описан у многих видов животных. Так, анализ θ-ритма кролика, кошки и обезьяны был проведен Грином и Ардуини (1953). Он описан у ежей, австралийского фалангера, крысы и морской свинки (Liberson, Akert, 1955; Weiss, Fifkova, I960; Adev et al. 1960; Oshima et al., 1962).
У человека выраженность θ-ритма в ЭЭГ зависит от возраста, фона основной активности, степени умственного напряжения. Признаком патологии, видимо, надо считать высокое содержание симметричных θ-волн, а также асимметричные и пароксизмальные θ-волны у взрослого человека в состоянии бодрствования. θ-ритм отражает вовлечение в патологический процесс, видимо, глубоких отделов мозга. Однако Е. А. Жирмунская (1959) полагает, что низкоамплитудный (25—35 мкВ) θ-ритм в ЭЭГ человека может составлять один из компонентов нормальной ЭЭГ. И. С. Берптов и др. (1943) считали, что медленные волны 4—6 кол/с в ЭЭГ здоровых людей являются слиянием 2—3 волн α-ритма; они обозначали их η-волнами. Эта точка зрения по нашла общего признания.
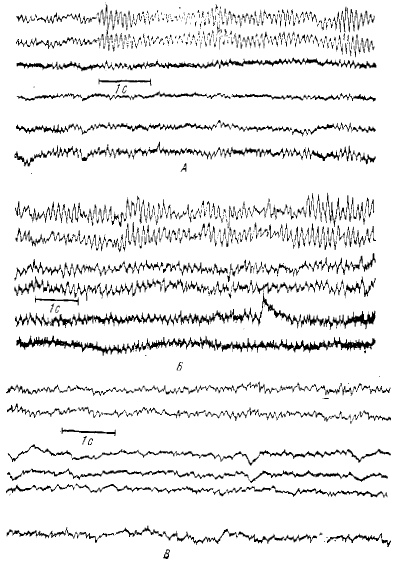
Альфа (α)-ритм состоит из волн частотой около 10 кол/с (от 8 до 13 кол/с) длительностью в среднем 90—120 мс. α-волны имеют «синусоидальную» форму и амплитуду порядка 20—60 мкВ (рис. 9). Могут, однако, наблюдаться высоко- и низкоамплитудные α-волны. Впервые термин α-ритм был введен, как мы уже говорили, Бергером в 1929 г., и ряд исследователей (например, Adrian, Maltews, 1934) назвали этот ритм бергеровским ритмом, α-ритм можно наблюдать во многих областях мозга человека. Он регистрируется как в обширных областях коры, так и во многих подкорковых образованиях— таламусе, базальных ядрах и др. (Okuma et al., 1954; Sem-Jacobsen ci al., 1955, 1956). Во всех точках коры мозга a-активность имеет одну и ту же частоту и постоянные фазовые соотношения.
α-ритм характерен для ЭЭГ взрослого здорового человека при состоянии бодрствования и закрытых глазах. Любая активация, усиление внимания и т. д. ведут к угнетению, блокаде α-ритма. При наркозе и естественном сне α-ритм также исчезает. Таким образом, α-ритм является своеобразной характеристикой определенного функционального состояния коры больших полушарий. Сдвиг в сторону от оптимального уровня — уровня α-активности (понижение или повышение степени активации) ведет к угнетению α-активности (Кратки, 1957). Если на фоне бодрого, спокойного состояния (когда в ЭЭГ хорошо выражен α-ритм) давать активирующие воздействия, наблюдается блокада α-ритма и те же воздействия на фоне дремотного состояния (когда α-ритм отсутствует) вызывают вспышку α-ритма. Частота α-ритма у разных испытуемых заметно отличается, по она крайне стабильна для каждого испытуемого (Брейзье. 1962), Обычно амплитуда α-волн испытывает определенную модуляцию: периодически то увеличивается, то ослабляется (сквик-эффект, по Уолтеру). В результате этого α-волны собираются в своеобразные веретена (см. рис. 9), длительность которых колеблется от 0,5 до 5 с. и более. В ряде случаев такое веретено α-ритма начинается с более высокочастотных волн, но через 0,5—1 с частота их снижается до обычной. Амплитуда α-ритма в разных точках может быть неодинакова.
Ранее α-ритм связывали преимущественно со зрительным анализатором, так как именно в затылочной области коры он имеет большую амплитуду и значительно легче блокируется световыми раздражениями. Предполагали, что именно здесь расположен первичный корковый очаг α-ритма, однако это не подтвердилось при более тщательных исследованиях.
Было показано (Garouttc et al., 1956, 1959), что могут быть различные случаи распределения α-ритма. В одних случаях изучение фазовых соотношений α-колебаний показало, что возможно постепенное волнообразное распространение α-волн от задних областей коры к передним, α-колебания в затылочной области коры возникали на 12—20 мс раньше, чем в центральных, и на 40—50 мс раньше, чем во фронтальных. Однако можно наблюдать и противоположный эффект — более раннее появление α-ритма в передних областях и почти так же часто, как и в затылочной области; причем α-волна из передней области может распространяться к затылку. Близкие данные получили и другие исследователи (Ю. В. Дубикайтис и В. В. Дубикайтис, 1962, 1963; Cooper, Mandy-Castle, 1960, и Др.). Это даст основание полагать, что α-ритм может первично возникать в разных зонах коры мозга человека, хотя фокус его активности привязан, как правило, к затылочной области коры. Последнее подтверждается исследованиями электрической активности мозга человека при нарушении зрения, которые показали прямую связь α-ритма со зрительной функцией.
Впервые Эдриан (Adrian, Mattews, 1934; Adrian, Jamagiva, 1935) высказал предположение о роли зрительной афферентации в формировании α-ритма. До последнего времени существовали разноречивые данные по этому вопросу. Данные, полученные в клинике человека, свидетельствовали об исчезновении α-ритма при различных повреждениях зрительного анализатора, вызывающих слепоту или значительную редукцию функции зрения. Ряд других исследователей полагают, что α-ритм прямо не зависит от зрительной афферентации и что он имеет место даже у слепых.
Л. А. Новикова (1966), подвергнув этот вопрос специальному экспериментальному анализу, пришла к заключению, что у слепых людей и особенно у слепорожденных α-ритм не регистрируется у людей с дефектом зрения по мере возрастания остроты зрения степень выраженности α-ритма на ЭЭГ увеличивается. Предпринятый Л. А. Новиковой анализ механизмов этого явления привел к заключению, что депрессивный характер ЭЭГ слепых и в том числе отсутствие α-ритма связано со снижением тонуса коры и обусловлено выключением ведущей в жизни человека зрительной афферентации.
В настоящее время различают α-активность и α-ритм. Первая обычно состоит из одиночных α-волн с длительностью от 80 до 125 мс. Эта α-активность регистрируется в любых мозговых структурах; α-ритм определяется как регулярная волновая активность с частотой порядка 10 Гц и регистрируется преимущественно в теменно-затылочных областях.
Из работ ряда авторов известно, что у человека к 13 годам электрическая активность мозга приобретает черты, свойственные ЭЭГ взрослых людей. ЭЭГ детей 10—12-летнего возраста по степени выраженности α-ритма в затылочных областях коры отличается от ЭЭГ взрослых людей незначительно, α-ритм остается четко выраженным до 50—60 лет. Однако в электроэнцефалографии детского возраста существуют до сих пор противоречия по вопросу характеристики отдельных компонентов ЭЭГ и их классификации (Фарбер, 1969). Ряд авторов полагают, что α-ритм формируется у ребенка из ритмических колебаний 3—4 в 1 с, преобладающих в ЭЭГ маленьких детей. Уолтер (Walter, 1953) и другие авторы считают, что увеличение частоты α-ритма с возрастом не происходит. Они полагают, что в процессе развития сменяются нейронные механизмы генерации ритмов, разных по частоте и своему генезу.
Многие авторы относят к α-ритму маленьких детей ритмы ЭЭГ 3—5 в 1 с, другие — 6—8 в 1 с, а третьи — 8—9 в 1 с. Другие категорически возражают против этого, полагая, что все эти колебания являются не генетическим аналогом «будущего» α-ритма, а представляют собой самостоятельные колебания другой природы.
Очевидно, что выявление удобной модели α-ритма У животных может сыграть важную роль для изучения его генеза (Гусельников, Супин, 1968). Активность, очень сходная с α-ритмом, наблюдается у обезьян (Cavennes, 1962), кошек (Clark, Ward, 1945; Крагин, 1962) и других животных. Описан и еще один вид α-подобной активности у кролика, которая, возможно, аналогична α-ритму человека. Это ритмические колебания потенциала в затылочной коре бодрствующего спокойного кролика, имеющие частоты 4—5 в 1 с (Bartley, 1940; Bishop, Clare, 1952, и др.). Эта активность имеет ряд свойств, сближающих ее с α-ритмом человека: фокус ее также локализован в затылочной коре в области проекции зрительного анализатора; этот ритм угнетается, как и α-ритм человека, при общей активности животного и при действии световых раздражителей. Все это дает основание считать, что он может являться аналогом α-ритма (Bartley, 1940).
Роландический ритм был открыт в 1952 г. Гасто и затем описан в работах других авторов (Gastaut et al., 1957; Klass, Bicford, 1957; Роже, Воронин, Соколов, 1958; Chatrian et al., 1959, и др.). Роландический ритм еще называют μ-ритмом, гребенчатым, дугообразным или аркообразным ритмом.
Этот ритм регистрируется с черепа или с обнаженной поверхности мозга только у человека з области роландовой борозды, где всегда находится его фокус. Он напоминает по своей частоте и форме α-ритм, но отличается отпето по ряду внешних признаков и по физиологическим свойствам. Его частота 7—11 кол/с. Форма веретенообразная, но часто может иметь вид однофазных колебаний характерной формы, отчего за ним в литературе закрепилось название «аркообразный», или «wicket»-ритм.
Роландический ритм в норме можно зарегистрировать лишь у небольшого числа испытуемых (например, у 15 из 500, т. е. в 3% случаев, Chatrian et al., 1959). Он отличается от α-ритма по ряду функциональных свойств. Если α-ритм наиболее чувствителен к световым раздражениям, то роландический ритм — к тактильным и проприоцептивным раздражениям, а также к воображаемым движениям. Депрессия ритма при всех этих раздражениях лучше всего выражена в противоположном полушарии, что говорит об особой роли проприоцептивной афферентации в его блокаде. Л. А. Новикова отмечает, что у слепых на общем фоне депрессии корковой ритмики происходит формирование роландического ритма, она связывает это с компенсаторным усилением деятельности двигательного анализатора.
Роландический ритм, вероятно, может быть идентифицирован с рядом описанные ранее вариантов α-ритма (Кожевников, Мещерский, 1963), прецентральным α-ритмом (Jasper, Andrews, 1938), высокоамплитудным α-ритмом (Schutz, Muller, 1951) и α-подобной активностью (Maddocks et al., 1951). Иногда его определяют как β-вариант-ритм, подчеркивая его функциональную и тонографическую общность с β-ритмом, что вряд ли оправдано.
Несмотря на различия роландического ритма и α-ритма, есть принципиальное сходство между ними: так же как и α-ритм, он наблюдается при неактивном, расслабленном состоянии двигательной системы, прямо зависит (блокируется) от активации ведущего для него двигательного анализатора, имеет близкую частоту с α-ритмом и т. д. Можно ожидать, что они действительно являются различными модификациями одного и того же процесса. Именно в связи с этим можно отметить, что при отведении от височных областей черепа человека у 50% испытуемых умственные нагрузки вызывают не депрессию, а появление в ЭЭГ ритмической веретенообразной активности с частотой (8—12 кол/с) и амплитудой 20—30 мкВ (Kennedy et al., 1948). Авторы обозначили волны этого ритма каппа (κ)-волны, а ритм — κ-ритм (или Кеннеди волны). Таким образом, и в этой области коры полушарий наблюдается активность, сходная с α-ритмом. Возможно, что все эти сходства являются отражением еще не ясных общих механизмов и общих принципов генеза всех этих ритмических колебаний коры, получающих лишь «специфическую окраску», в основе которой лежит та или иная модальность афферентного потока. Что касается гомологов роландического ритма у животных, то некоторые исследователи считают таковым «сенсомоторный» ритм в коре у кошки (Roth et ah, 1967; Chase, Harper, 1971; Howe, Sterman, 1972; Rougeul et al., 1972).
Бета (β)-ритм коры человека состоит из быстрых волн длительностью до 40—50 мс и частотой 14—30 кол/с (см. рис. 9). Впервые термин β-ритм был введен Бергером. Обычно β-ритм связывают с сенсомоторной системой.
И. С. Беритов (1948) полагает, что этот ритм коры определяется деятельностью нервных кругов и цепочек коры, что наблюдается обычно при активном состоянии ретикулярной формации ствола мозга; при ее блокаде β-ритм исчезает.
У большинства животных в разных отделах мозга, в том числе и в коре больших полушарий, можно зарегистрировать сходные и более высокочастотные ритмы. Ряд исследователей их обычно не классифицируют как β-ритм, оставляя этот термин лишь для ЭЭГ человека, тем более что об общности их генеза ничего не известно.
Однако И. С. Беритов (1969) относит к β-активности высокочастотные ритмы в ЭЭГ кошки (до 500 в 1 с) и в ЭЭГ обезьян (до 200—300 в 1 с). У человека к β-активности он относит волны с частотой от 250 до 480 в 1 с. Чем выше частота β-ритма, тем ниже его амплитуда.
β-ритм человека регистрируется, как правило, в лобных отделах мозга. Генерализованная по коре низкоамплитудная быстрая β-активность является показателем патологии. Так, Флюгель (Fliigel, 1968) отмечает, что в группе больных с высокочастотными β-волнами (больше 30 в 1 с) отмечен большой процент психовегетативных синдромов.
Оценивая функциональное значение β-ритма на разных этапах развития патологического процесса при эпилепсии, опухоли мозга и травме, И. В. Введенская и др. (1971) рассматривают его как показатель включения или усиления структур мозга, участвующих в компенсаторных процессах. Ими показано, что наличие β-ритма в ЭЭГ больных в восстановительном периоде после травмы черепа и мозга имеет благоприятное прогностическое значение,
Гамма (γ)-ритм состоит из волн низкой амплитуды не более 15 мкВ, имеющих частоту 30—35 кол/с. Он выражен лучше в передних отделах мозга. Если его амплитуда выше 15 мкВ, то ЭЭГ рассматривается как патологическая (Кожевников, Мещерский, 1963).
Сигма (σ)-ритм впервые для человека был описан Лумисом с соавторами (Loomis et al., 1935) при исследовании электроэнцефалографического выражения различных стадий сна.
Моменты раздражения отмечены стрелками, артефактами и точками
σ-ритм является одним из основных и наиболее четко выраженных компонентов спонтанной ЭЭГ млекопитающих, находящихся в состоянии естественного сна или сна подобных состояниях, вызванных некоторыми фармакологическими и нейрохирургическими вмешательствами. Большинство исследователей считают, что σ-ритм появляется в ЭЭГ человека при наступлении легкого сна (Gibbs, 1950; Dement, Kleitman, 1957; Hess, 1964, и др.). Спонтанный σ-ритм представляет собой вспышки ритмических электрических колебаний с частотой до 10—16 в в 1 с. Появление в виде отдельных групп волн с постепенно нарастающей и затем убывающей амплитудой (рис. 10) является характерным внешним признаком σ-ритма, за что он и получил другие названия — веретенообразная активность (spindle activity), взрывная или вспышечная активность (burst activity), веретенообразные вспышки (spindle burst), α-подобный ритм животных, сонные веретена (sleep spindles), барбитуровые веретена (barbiturate spindles) и ряд других. Надо полагать, что термины веретена и вспышки должны быть сохранены за целым классом ЭЭГ колебаний, имеющих характерные амплитудные модуляции (например, за α-ритмом, роландическим ритмом, обонятельным ритмом) и обозначение этим термином α-ритма— не целесообразно. Термин α-подобные также неприемлем, т. к. α-ритм и α-ритм — совершенно различные виды электрической активности мозга (Гусельников, Мухаметов, 1967, 1969). Название сонные веретена и барбитуровые веретена удобно сохранить за разновидностями этого ритма. Мы и дальше, будем все его модификации обозначать σ-ритмом. Спонтанные его проявления будем называть σ-веретенами, а вызванные одиночным раздражением мозга — σ-послеразрядами. За некоторыми разновидностями σ-ритма, видимо, рационально сохранить наиболее привычное обозначение. К спонтанному σ-ритму относятся σ-веретена естественного сна (сонные веретена), наркотического сна. вызванного барбитуратами (барбитуровые веретена), препаратов мозга с различными хирургическими нарушениями активирующих систем и при некоторых других состояниях мозга.
Вызванный σ-ритм может быть получен либо одиночным возбуждением механизма этого ритма (σ-послеразряды), либо ритмическим возбуждением на резонансной частоте 10—14 в 1 с.
К σ-послеразрядам относятся специфические σ-послеразряды, которые возникают на таламо корковом уровне специфических афферентных систем в ответ на короткий адекватный стимул или одиночное электрическое раздражение любого уровня данной системы (рис. 10). Многие авторы (Dempsey, Morison, 1943; Chang, 1950, Bremer, 1958, Andersen, Eccles, 1962) называют эти ритмические колебания по-разному.
Под специфическими послеразрядами мы будем подразумевать послеразряды, возникающие только в слуховой и соматической афферентных системах. Кроме того, σ-послеразряды могут быть вызваны коротким электрическим раздражением неспецифического таламуса, неокортекса, хвостатого ядра (веретена хвостатого ядра — caudate spindles), мезенцефалической ретикулярной формации и некоторых других структур (см. рис. 10). Необходимо отметить, что при ритмическом возбуждении механизма σ-ритма возникает реакция вовлечения, если раздражается неспецифический таламус при связанные с ним структуры, или реакция усиления, если раздражается специфический таламус. В соответствии с этим целый ряд спонтанных и вызванных ритмических процессов, часто изучавшихся изолированно, имеет, как мы убедимся, по существу, одни и тот же механизм и представляет собой модификацию одного и того же ритма.

А — Обонятельная луковица (ОЛ) лягушки (хронический опыт); Б— гиппокампальная кора (1) и ОЛ (2) черепами. В - OЛ кролика
Обонятельный ритм. Под обонятельным ритмом подразумевается ритмическая электрическая активность обонятельного анализатора животных и человека, впервые описанная у животных Эдрианом (Adrian, 1942, 1956) и названная им обонятельными вызванными волнами.
Обонятельные вызванные волны в обонятельной луковице (ОЛ) и ряде других образовании мозга (в основном, структурах обонятельного мозга) регистрируются у человека и позвоночных животных всех классов (рис. 11 и табл. 2), В отличие от млекопитающих и человека, у рыб, амфибий, рептилий и птиц они являются, видимо, единственным видом королю выраженной ритмической электрической активности мозга (Гусельников, 1965; Гусельникова, Гусельников, 1975). Вероятно, незначительные изменения структурных особенностей обонятельной луковицы и ее связей в ряду позвоночных свидетельствуют о том, что вызванные волны имеют одинаковую природу у представителей различных классов и видов позвоночных. Такое предположение в некоторой степени подтверждается сходством электрических реакции обонятельной луковицы у различных классов позвоночных, а также отсутствием заметных их различий у макро- и микросматиков (Oltoson, 1963).
Таблица 2. Частота вызванных волн в обонятельной луковице позвоночных
| Объект исследования | Частота волн, кол/с | Авторы | ||
|---|---|---|---|---|
| Рыбы | 6—8 | Гусельникова, Гусельников, 1975 | ||
| Амфибии | 6—9 | Geets, 1948; Takagi, Shibuya 1960 | ||
| Рептилии | 15—25 | Winkel, Caspers, 1953; Гусельников, 1956 | ||
| Птицы | 15—25 | Гусельников, 1964 | ||
| Млекопитающие | ||||
| Насекомоядные (еж) | 15—45 | Adrian, 1942 | ||
| Грызуны (кролик) | 55 | Гедеванишвили, 1955 | ||
| Кошка | 45 | Adrian, 1942 | ||
| Собака | 20—40 | Ueki, Domino, 1961 | ||
| Обезьяны | 40 | Hughes, Mazurowski, 1962; Ueki, Domino, 1961 | ||
| Человек | 38—70 | Гедеванишвили, 1955; Sem — Jacobsen et al., 1956 | ||
Вспышка вызванных обонятельных волн имеет форму веретена и обычно появляется, в фазе вдоха, хотя их можно наблюдать и в фазе выдоха (Ueki, Domino, 1961). Обонятельные вызванные волны можно получить в остром опыте у всех видов животных «протягиванием» воздуха через носовую полость трахеотомированного животного (Новикова, Хволес, 1953; Гусельников, 1964; Adrian, 1942, и др.). Отсюда обонятельный ритм и получил название «обонятельные вызванные волны», хотя он регистрируется спонтанно во время вдоха или выдоха животного или особенно во время ориентировочной реакции «принюхивания».
У представителей различных классов позвоночных частота волн во вспышке разная. В ряду рыбы — амфибии—рептилии—млекопитающие наблюдается явная тенденция к увеличению частоты. Что касается ритмической обонятельной активности, регистрируемой в других структурах мозга, с которыми обонятельная луковица имеет прямые связи, то было показано, что у кролика (Новикова, Хвотес, 1953) обонятельные вызванные волны могут иметь место на всем пути oт обонятельной луковицы вплоть до переднего гипоталамуса. У кошек эти волны имеют место в амигдало-препириформной области и гиппокампе (Gault, Coustau, 1965), у обезьян и собак в обонятельной полоске, пириформной и препириформной коре, медиальной миндалине., заднем гипоталамусе (Domino, Ueki, 1960), у крысы — в препириформной коре (Гусельников, 1964), у ящерицы (Winkel, Caspers, 1953) и черепахи (Гусельников, 1956, 1964) в гиппокампальной коре и у птиц в septum, в медиальном гиперстриатуме (Гусельников, 1964) и т. д.
Таким образом, почти у всех представителей различных классов позвоночных в центральных структурах обонятельного анализатора возникают обонятельные волны, сходные по конфигурации и частоте с вызванными волнами обонятельной луковицы.
Спайк-волна ритм. Характеристики этого ритма мы уже касались ранее, при рассмотрении комплекса спайк-волна.
Дыхательный ритм. При регистрации ЭЭГ могут наблюдаться артефакты, связанные с ритмом дыхания. Обычно это связано с механическим смещением отводящих электродов на коже черепа в момент вдоха и выдоха.
А — дыхательный pитм и ЭЭГ кролика (по Бут и Климовой-Черкасовой, 1969): 1—3 —одномоментная запись по трем каналам электрической активности (1,2) и дыхания (3), 4 — время 1/4 с. Стрихнинные разряды в ЭГ переднего мозга ящерицы и голубя (по Гусельникову, 1960). 6 — ЭЭГ переднего мозга ящерицы и Г — голубя до — и В — ЭГ ящерицы и Д — голубя через 10—13 мин после отравления участка переднего мозга 0,1%-ным раствором стрихнина. Видны высоковольтные стрихнинные разряды, идущие по ритму дыхания, 1 — ЭГ полушарий переднего мозга, 2 — запись дыхания
Однако в ЭЭГ имеет место и физиологический дыхательный ритм — S-ритм (от слова «spiro» — дышу) (рис. 12). Этот ритм не является артефактом, но изучен пока крайне недостаточно. Обычно он регистрируется при специальных условиях (Ливанов, Поляков, 1945; Бут, Климова-Черкасова, 1969, и др.). Считается, что S-ритм в ЭЭГ проявляется при ухудшении функционального состояния коры (Григорьев, 1958; Гурвич, 1966; Бут, Климова-Черкасова, 1969, и др.). Так, Гурвич (1966) показал, что этот ритм регистрируется особенно четко при умирании организма.
Однако определенная зависимость синхронизации ЭЭГ здорового человека от дыхательного цикла была найдена рядом исследователей (Roth, Klingberg, 1970; Faber et a]., 1970, и др.).
Еще в 1936 г. А. И. Смирновым, а позднее М. В. Сергиевским (1950) и др. было показано влияние дыхательного центра на кору больших полушарий. В 1954 г. Ройтбак получил прямые доказательства иррадиации импульсов из дыхательного центра на кору больших полушарий. Он обнаружил, что судорожные потенциалы в отравленном стрихнином участке коры наступают точно по ритму дыхания. В наших экспериментах с использованием той же методики (Гусельников, 1960) был показан универсальный характер влияния дыхательного центра на вышележащие отделы мозга у позвоночных разного уровня эволюционного развития (см. рис. 12).
Источником S-ритма в ЭЭГ являются каудальные отделы ствола мозга, и большинство исследователей связывают его с дыхательным центром (Григорьев, 1958; Королькова, 1966; Латаш, 1959; Бут и Климова-Черкасова, 1969, и др.). Бут и Климова-Черкасова полагают, что S-ритм связан с распространением к коре импульсации по неспецифическим афферентным путям из ретикулярной формации каудального отдела ствола мозга через внутреннюю капсулу (экстраталамически).
Надо полагать, что восходящее тоническое влияние дыхательного центра связано с облегчающим действием на корковые нейроны (Угрюмов, 1947; Sharpless, Jasper, 1956) и имеет, видимо, постоянный характер, но проявляется в ЭЭГ при разных сдвигах функционального состояния мозга, в том числе и при ориентировочной реакции (Ливанов, Поляков, 1915; Бут и Климова-Черкасова, 1969). Специальный анализ связи ЭЭГ с дыхательным ритмом провел А. А. Генкин (1971). Он пришел к выводу не только о наличии такой связи, но и предполагает наличие локальных влияний дыхательного центра на определенные области коры. Тонические влияния дыхательного центра имеют место не только на уровне коры, но и на уровне, спинного мозга (Орбели, Кунстман, 1961; Латаш, 1959), а также таламуса и гипоталамуса (Бут, Климова-Черкасова, 1969).
Нерегулярная, или аритмическая, фоновая активность головного мозга
Обычно она состоит из волн самой различной длительности и амплитуды. Могут быть разные, случаи проявления этой активности. Так, у многих животных как в коре полушарий, так и в других отделах мозга нерегулярная, аритмичная активность может составлять основной рисунок ЭЭГ и уступать место ритмической, регулярной активности лишь при определенных физиологических состояниях (стресс-состояние, ориентировочная реакция, сон и т. д.).
В других случаях эта активность может наблюдаться в «интервалах» между различными «вспышками» той или иной регулярней активности. Провести какие-либо градации этой активности сложно, тем более что она, по сути дела, является суммой самых различных колебаний и в каждом отдельном случае необходим специальный анализ составляющих ее компонентов.
Пароксизмальная активность
➥ Основная статья: Пароксизмальная активность мозга
Обозначения местоположения регисрирующих электродов см. на рис. 7
Включает четко ограниченные группы высокоамплитудных волн, которые обычно возникают и резко выделяются на фоне более низкоамплитудного основного ритма (Кожевников, Мещерский, 1963). Так, пароксизмальные вспышки ритмов с частотой 10 и 25 кол/с могут наблюдаться в нормальной ЭЭГ при состоянии бодрствования, а вспышки 14 кол/с — при состоянии сна (вспышки σ-веретен) (рис. 13). У больных эпилепсией могут появляться самые различные пароксизмальные ритмы — комплексы спайк-волн, вспышки медленных волн и т. д. Пароксизмальные, ритмы, характеризующиеся фиксированной частотой, возникают внезапно, прогрессивно нарастают до максимума и внезапно прекращаются.
Продолжая дальше классификацию электрических процессов мозга, необходимо рассмотрев изменения фоновой ритмики (см. схему).
Изменения фоновой активности структур головного мозга животных и человека “возникают “при самых различных условиях: действии внешних раздражителей, естественных состояниях сна и бодрствования, ориентировочной и условнорефлекторной деятельности, течении сложных безусловных реакций, раздражении мозговых структур, действии различных физических и гуморальных факторов и т. д. Ряд этих воздействий вызывает общие сдвиги текущей спонтанной активности мозга, часто не равнозначные в различных мозговых структурах.
Поэтому, чтобы в дальнейшем легче было разобраться в генезе и динамике изменений фоновой активности, отметим несколько групп общего типа реакций, имеющих место в коре больших полушарий животных и человека:
- Реакцию активации;
- Реакцию инактивации;
- Искусственные способы изменения фоновой ритмики.
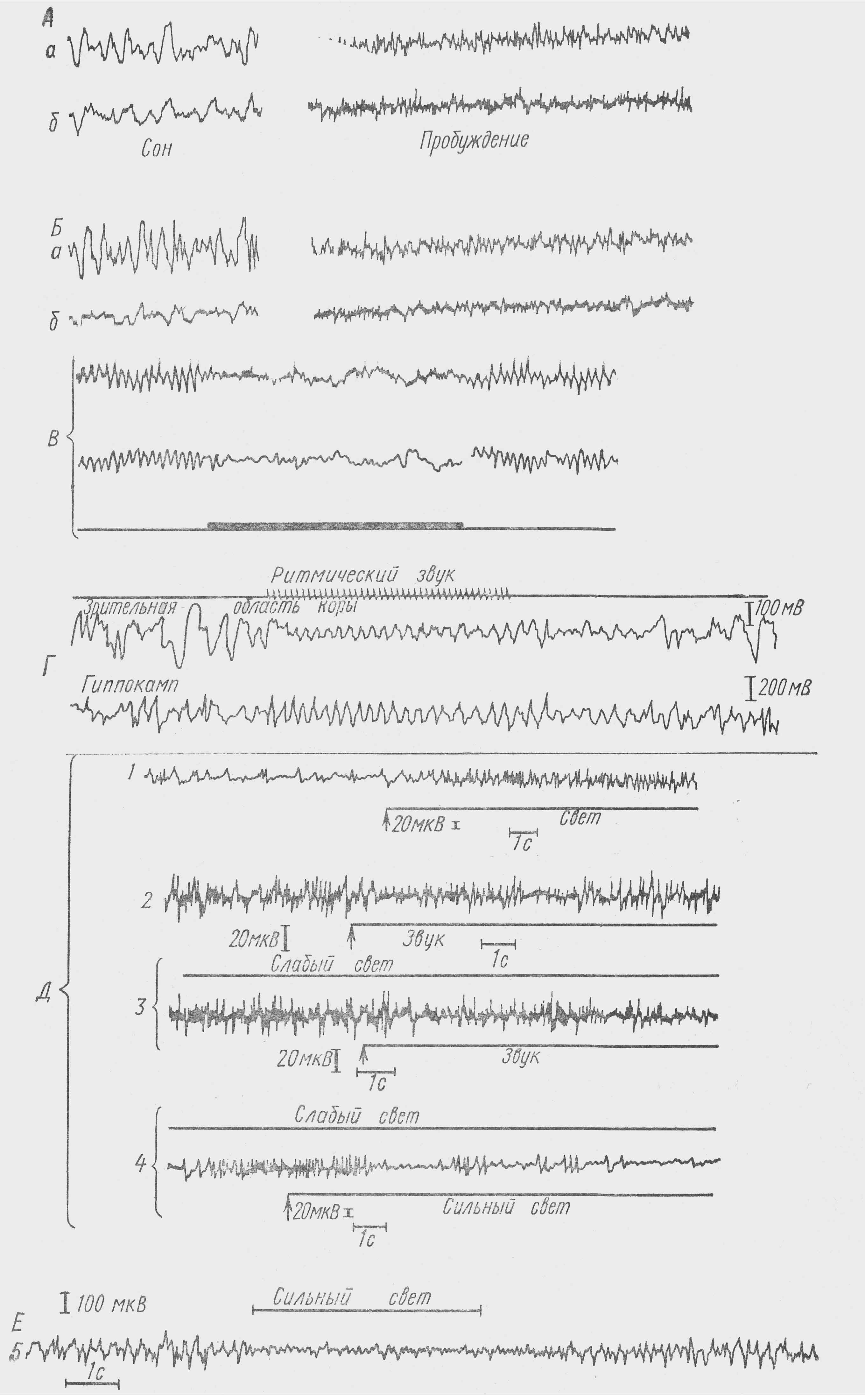
1 — усиление активности коры среднего мозга ящерицы при действии света на фоне низкой спонтанной активности, 2 — реакция де¬прессии в коре ящерицы при действии звука на фоне высокой спонтанной активности, 3 — при действии звука и 4 — действии силь¬ного света при предварительной активации слабым светом, 5 — реакция депрессии активности в ЭГ полушарий переднего мозга
Реакция активации. Еще Бергер (Berger, 1929) отметил, что открывание глаз, внешние раздражения, умственное напряжение приводит к депрессии, угнетению, блокаде α-ритма. В 1937 г. Рейнбергер и Джаспер (Reinberger, Jasper), исследуя ЭЭГ и поведение котики, наблюдали пробуждение животного при действии разных афферентных раздражений. Они выяснили, что все применяемые раздражения, вызывающие пробуждение и бодрствование, сопровождались сменой в ЭЭГ медленных ритмов на низковольтную быструю активность. Эта реакция была названа ими реакцией активации. Они же отметили генерализованный характер этой реакции. Позже Моруцци и Мэгун показали, что прямое раздражение ретикулярной формации ствола мозга воспроизводит все изменений, характерные для активации, связанной с бодрствованием, ничем не отличающейся от реакции при действии внешних раздражителей. С этого момента реакцию угнетения медленной ритмики, или реакцию активации, начинают чаще называть реакцией пробуждения, или эраузал-реакцией (arousal reaction).(см. рис. 14)
Часто применяется и другой термин для этой реакции— десинхронизация. Этот термин не точен и не применим для всех случаев реакции активации. В относительной степени он может быть использован в реакции, когда четкая ритмическая (синхронизированная) активность мозга сменяется высокочастотной активностью. Мы будем пользоваться термином реакция активации.
У человека (Соколов, 1962) в зависимости от исходного фона активация с ЭЭГ проявляется: а) в усилении быстрых колебаний на фоне уже развившейся депрессии α-ритма, б) постепенной депрессией α-ритма и увеличением доли в суммарной ЭЭГ β- и γ-колебаний. в) депрессией медленных δ и θ-колебаний и сменой их или высокоамплитудным α-ритмом, или же повышением β-активности.
Реакции активации противопоставляют реакцию инактивации (Соколов, 1962), которая связана со снижением уровня возбуждения в коре. Эта реакция, в зависимости от исходного уровня активности, выражается в появлении α-ритма вместо быстрой низкоамплитудной активности, появлении медленных δ и θ-волн вместо β- и α-колебаний, дальнейшем замедлении и возрастании амплитуды медленных волн, уже присутствующих в фоновой активности.
Изменение ритмики электрических колебаний коры, как в случае реакции активации, так и при инактивации, может быть различной длительности и носить фазический и тонический характер (Соколов, 1956). По данным Шарплесса и Джаспера (Sharpless, Jasper, 1956), тоническая реакция связана с участием стволовой части ретикулярной системы, а фазическая — с неспецифической системой таламуса.
Обычно реакция активации сопровождает ориентировочную деятельность человека и животных, и она, как мы указывали, как правило, связана с генерализованным по коре угнетением спонтанной ритмики и появлением быстрой, высокочастотной активности. Однако у ряда животных ориентировочная реакция сопровождается появлением синхронизированных ритмов в ЭЭГ. Так, у кроликов и крыс в ответ на афферентные раздражения (звук, свет и т. д.) возникают синхронизированные колебания с частотой 4—7 в 1 с (Green, Arduini, 1954; Соколова, 1954; Новикова, Фарбер, 1959; Семпохина, 1958; Гусельникова, 1958; Чжоу-Шао-Цы, 1958; Банцекипа, 1962, и др.). Особенно четко эти синхронизированные ритмы при ориентированной реакции наблюдаются в древних структурах мозга — гиппокампе, гипоталамусе и ретикулярной формации.
Систематические исследования, проведенные нами (Гусельников, 1964) на рыбах, амфибиях, рептилиях и птицах, показали, что у хронически оперированных животных с вживленными электродами ориентировочная реакция сопровождается в электрограмме различных отделов мозга сложным характером изменении суммарной активности, который зависит от многих факторов, и прежде всего от исходного фона активности, который в свою очередь определяется внешними условиями. В связи с этим реакция может выражаться либо в угнетении, либо в усилении фоновой активности, что, видимо, связано со спецификой развития в филогенезе функций ретикулярной формации ствола мозга.
Искусственные способы изменения фоновой ритмики
Включают изменения активности мозговых структур, вызванные прямым действием ряда химических и физических факторов: локальная аппликация на поверхность мозга различных фармакологических препаратов, прямое электрическое раздражение, анодная и катодная поляризация нервных структур, прямое действие механических и физических факторов и т. п. Все эти воздействия входят в арсенал средств экспериментальной нейрофизиологии, используемых для анализа механизмов электрогенеза суммарной активности мозга. Также воздействия могут вызвать изменения фоновой активности, не имеющие места в естественных условиях. Так, например, при аллокации кусочка фильтровальной бумаги, смоченной 2—2,5%-ным КСl, на поверхность мозга возникает вначале локальное, а затем распространяющееся по коре полное подавление электрической активности. Это явление получило название по имени автора — распространяющаяся депрессия Лео (Leao, 1944). Распространяющаяся депрессия как прием исследования была широко использована для решения ряда интересных вопросов нейрофизиологии коры. Этот и ряд других приемов искусственного воздействия на фоновую активность мы встретим в разных разделах нашего курса.
Длительные электрические (сверхмедленные) колебания потенциала
Наличие медленных длительных потенциалов в коре головного мозга было показано многими авторами (Русинов, 1947, 1962; Швец, 1958; Аладжалова, 1960, 1962; Brookhart et al., 1958, и многие другие). Значительно сложнее и запутанней обстоит дело с классификацией всех этих медленных и сверхмедленных колебаний потенциала. Эти потенциалы называют: апериодические медленные колебания, медленные длительные потенциалы, изменения уровня постоянного потенциала, сверхмедленные колебания, гладкие потенциалы, квази- постоянный потенциал и т. д.
В 1969 г. Н. А. Аладжаловой была предложена классификация длительных потенциалов, которой мы будем придерживаться. По характеру проявления она предложила разделить их на:
- Постоянные (стабильные) потенциалы;
- Апериодические сдвиги постоянного потенциала;
- Ритмические (сверхмедленные) колебания постоянного потенциала. По временному параметру последние делятся на секундные, минутные, многоминутные и часовые.
В целом колебательные потенциалы с такими временными периодами следует определять как сверхмедленные в отличие от δ и θ-ритма ЭЭГ. К числу длительных потенциалов головного мозга следует отнести и потенциалы, регистрируемые в коре больших полушарий при прямом электрическом ее раздражении.
Трудности, связанные с классификацией длительных потенциалов головного мозга, определяются прежде всего тем, что их происхождение остается до сих пор не совсем ясным; имеется много гипотез на этот счет, но ни одна из них, как отметил Ройтбак (1969), не является еще фактически достаточно обоснованней. Остается еще далеко неясным и вопрос о функциональном значении этих потенциалов, хотя определенная связь, например сдвига постоянного потенциала с изменениями ряда других суммарных реакции мозга, говорит об определенной их роли в интегративных функциях нервной системы.
Постоянные (стабильные) потенциалы регистрируются при отведении с разных участков мозга. Между поверхностью коры и белым веществом, а также между поверхностью коры и спинномозговой жидкостью и кровяным руслом имеет место постоянная разность потенциалов. При естественном сие и при наркозе потенциал коры смещается в положительную сторону, при бодрствовании— в отрицательную.
Апериодические сдвиги постоянного потенциала наступают при искусственных раздражениях различных нервных структур, в том числе и коры, адекватной или электрической стимуляции рецепторов, воздействиях на метаболизм мозга, при безусловной и условнорефлекторной деятельности, изменений кровообращения и аппликации различных веществ. Эти сдвиги постоянного потенциала (или, как еще говорят, сдвиги уровня постоянного потенциала, Русинов, 1969) могут носить локальный или диффузный характер, что зависит, видимо, от условий опыта (применяемых электродов и способов отведения, уровня наркоза, функционального состояния животного и т, д.), а также от характера применяемой стимуляции.

Записи сделаны при разных физиологических состояниях
Сверхмедленные ритмические колебания потенциала — колебания, близкие по форме к синусоидальным, с частотой 6—8 в 1 мин (при амплитуде 0,3—0,8 мВ) — или секундный ритм (рис. 15, А) и колебания с частотой 0,5— 2 в 1 мин (при амплитуде 0,5—1,5 мВ) —минутный ритм (рис. 15, Б). В разных образованиях головного мозга эти колебания проявляются различно. Наблюдаются они преимущественно в коре больших полушарий и в гипоталамусе. В ретикулярной формации ствола и таламуса они не найдены.
Длительные потенциалы коры на ее прямое электрическое раздражение
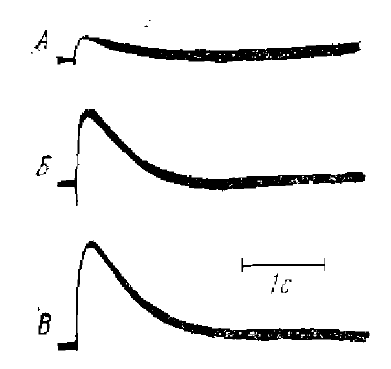
Расстояние между раздражаемым пунктом и «активным» отводящим электродом 0,75 мм
В ответ на относительно сильный стимул, нанесенный на поверхность коры, после начального отрицательного потенциала длительностью 20— 30 мс, возникает медленный отрицательный потенциал (МОП), продолжительность которого равна сотням миллисекунд и даже секунд (Ройтбак, 1965). МОП возникает при напряжении стимула в десять раз большем, чем пороговый стимул для вызова дендритного потенциала (Ройтбак, 1969). Он достигает амплитуды 2—3 мВ и продолжительности 0,5—3 с; латентный период равен 15—20 мс. Регистрируется до 3 мм от пункта раздражения только в верхних слоях коры (рис. 16). При ритмических электрических раздражениях коры возникает постоянное отрицательное отклонение, которое, как считает Ройтбак, является результатом суммации медленных отрицательных потенциалов, МОП используется в экспериментальной ЭЭГ для выяснения некоторых особенностей коркового электрогенеза.