Сейчас не вызывает сомнения тот факт, что нейрофизиологические механизмы, лежащие в основе σ-ритма, являются основой и целого ряда других ритмических процессов мозга, возникающих при сенсорном, электрическом и фармакологическом его раздражении, а также при нейрохирургических воздействиях.
К ним прежде всего относятся: реакция вовлечения (РВ) и реакция усиления (РУ), барбитуровые веретена наркотического сна, σ-веретена препарата cerveau isole, ряд σ-послеразрядов, возникающих при электрическом раздражении хвостатого ядра, специфических и неспецифических ядер таламуса и при сенсорных адекватных раздражениях.
➥ Более подробно: Общая классификация различных видов электрической активности головного мозга и их краткая характеристика
Все эти феномены в ЭЭГ животных по своим главным характеристикам крайне сходны. Это касается их частотного диапазона, характерной для них веретенообразности амплитудных модуляций волн, синаптических процессов в таламических нейронах, первично-таламического происхождения и т. д.
Видимо, более логично начать изложение данных о генезе σ-ритма с некоторых общих данных о РВ и РУ, а затем уже перейти к общим механизмам σ-ритма, где эти реакции будут рассматриваться Лишь как частный случай его проявления.
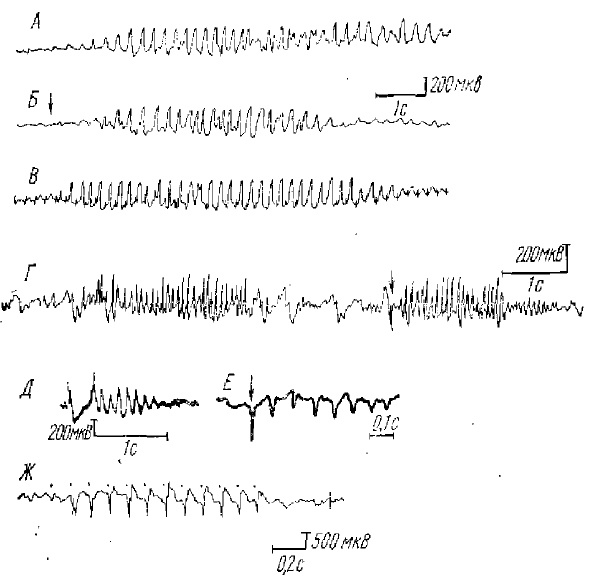
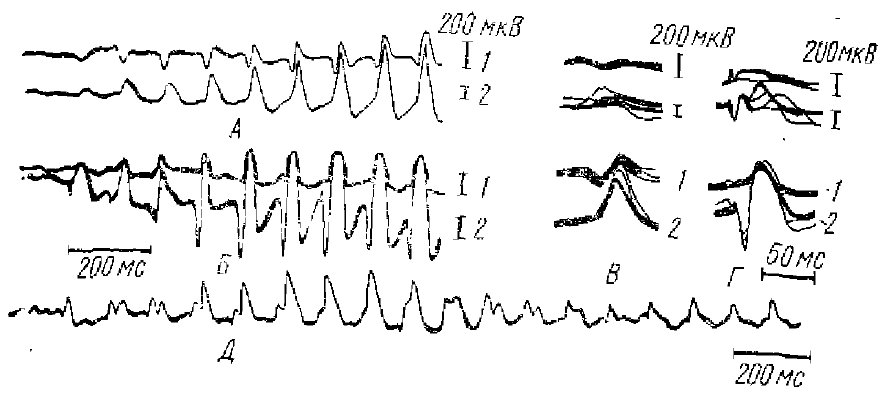
Впервые РВ, как мы уже отмечали, была описана Морисоном и Дэмпси (Morison, Dempsey, 1942) при ритмическом низкочастотном раздражении срединных ядер таламуса у кошек. При этом на каждый стимул в коре возникает электрический ответ. Эти ответы были длительными (20—35 мс), имели на поверхности коры негативную полярность и обладали характерным свойством «вовлечения», «рекрутирования» — амплитуда ответов при включении стимуляции постепенно нарастала от стимула к стимулу, достигая максимальной величины (намного превосходящей величину ответа на одиночное или первое из серии раздражение) только после подачи нескольких ритмических стимулов с частотой около 8— 12 имп/с (иногда от 5 до 15 имп/с).
РВ в коре мозга может возникать при раздражении практически всех ядер неспецифическои системы таламуса.
Вышеназванные авторы нашли, что по ряду признаков РВ обнаруживает сходство со спонтанной «взрывной», т. е. веретенообразной, активностью. А именно РВ и спонтанная веретенообразная активность имеют практически одинаковое распространение по коре мозга. Частота стимуляции, при которой легче всего вызвать РВ, очень близка к частоте спонтанной веретенообразной активности (в основном 8—12 Гц), величина начального эффекта вовлечения зависела от фазы спонтанной вспышки, на которой начиналась реакция, что указывает также на тесную связь между этими формами активности. При продолжительной ритмической стимуляции срединного таламуса корковые потенциалы РВ, достигнув после нескольких стимулов максимальной величины, не оставались на этом уровне, а обнаруживали периодическое увеличение и уменьшение амплитуды (фазы нарастания- спада), так что огибающая потенциалов РВ также имела веретенообразную форму, очень сходную с формой спонтанных веретен; оба вида активности регистрировались в участке коры, изолированной хирургически от остальной коры, по сохранившей связи с таламусом. Наконец, о тесной связи РВ со спонтанной веретенообразной активностью свидетельствуют исследования взаимодействия между собой этих двух видов активности, потенциалы РВ, возникающие во время развития фоновых волн веретенообразной активности, блокируются, что указывает на возможное участие одних и тех же элементов в генерации как фоновых веретен, так и РВ.
Моменты раздражения отмечены стрелками, артефактами и точками
Позже Дэмпси и Морисоном (Dempsey, Morison, 1943) было показано, что явление, сходное с РВ, может наблюдаться также и при ритмическом раздражении специфических, релейных ядер таламуса. В этих условиях (при ритмическом раздражении с частотой порядка 8 имп/с) к первичному потенциалу коры, возникающему в ответ на каждый стимул из ритмической серии, добавляется дополнительное колебание потенциала, обнаруживающее такую же способность к постепенному увеличению, рекрутированию по ходу ритмической стимуляции, как и потенциалы РВ. При одиночных раздражениях это дополнительное колебание не выявляется. Этот феномен и был назван Дэмпси и Морисоном реакцией усиления (РУ) (см. рис. 1). Было обнаружено, что РУ также обладает свойством взаимной блокады с фоновой биоэлектрической активностью, возникающей в специфической проекционной таламо-кортикальной системе.
Важные доказательства близости механизмов спонтанной веретенообразной активности и ритмических реакций на раздражение таламуса получили Спенсер и Брукхарт (Spencer, Brookhart, 1961).
Исследуя характер потенциалов спонтанных веретен, РВ и РУ, регистрируемых как от поверхности коры, так и от глубоких корковых слоев, они показали, что спонтанные ритмические потенциалы веретенообразной активности обнаруживают полное сходство с потенциалами РВ и РУ: часть волн спонтанных веретен имеют такую же форму и распределение потенциалов, по поперечнику коры, как волны РУ, а часть — как волны РВ (рис. 2, 3). Импульсная активность нейронов, сопровождающая РВ и РУ, также сходна с активностью во время спонтанных веретенообразных вспышек (рис. 4).
Все это даст основание считать, что РВ и РУ являются искусственно вызываемыми аналогами веретенообразной активности, т. е. ритмические раздражения таламических ядер активируют тот же механизм, который может генерировать и спонтанную активность типа веретен.
Представление о тесном сходстве РВ и РУ со спонтанной веретенообразной активностью прочно вошло в современную физиологическую литературу, вследствие чего эти реакции, очень удобные для экспериментального изучения, широко используются многими авторами в качестве типичных примеров ритмической электрической активности головного мозга — σ-ритма.
Общая характеристика σ-ритма (сонных веретен)
Сонные веретена описаны только у представителей различных отрядов млекопитающих; сумчатых, ежа, крота, броненосца, крысы, кролика, кошки, собаки, низших и высших обезьян и у человека.
Веретена отсутствуют у рыб, амфибий, рептилий и птиц (Гусельников, 1965; Mollica, Orsini 1958; Traclardi, 1966; Ookawa Takagi, 1968) и у однопроходных (ехидны). Несколько более спорен вопрос о наличии сонных веретен в коре рептилий, так как их легко спутать с характерными вспышками веретен обонятельного ритма, который хорошо выражен в общей коре у этих животных (Гусельников, 1965). Однако некоторые исследователи (Белехова, 1968; Servit, Strejckova, 1972) считают, что у рептилий они имеют место. Таким образом, в филогенезе позвоночных четко выраженный σ-ритм возникает, видимо, только у млекопитающих. Что касается онтогенеза, то описываются различные сроки появления σ-ритма. У грызунов он появляется на 10—15 день постнатального онтогенеза. К этому сроку хорошо выражена (например, у кролика) и РВ. У котят и щенков веретена появляются несколько позже. У человека сонные веретена возникают примерно с трехмесячного возраста. Частота их с возрастом не меняется.
У животных и человека сонные веретена возникают, как правило, в фазу медленного сна, следующего сразу за дремотой. Во время стадии сна, характеризующегося — δ-волнами, они возникают реже или полностью отсутствуют. При переходе к быстрому (парадоксальному) сну они могут вновь быть выражены в ЭЭГ, но полностью блокируются (как и РВ) в развитой фазе быстрого сна (Rossi, Favale, 1961; Giaquinto. 1968; Mancia et al., 1971). Спонтанные веретена регистрируются во время зимней спячки (Штарк, 1970).
Спонтанные веретена характерны для ЭЭГ при различных хирургических воздействиях, приводящих к снижению тонуса активирующих структур мозга. Так, в зависимости от уровня перерезки ствола мозга и состояния препарата веретена могут доминировать в ЭЭГ (препарат cerveau isole) и полностью отсутствовать при мостовом претригеминальном препарате. В хроническом опыте оба препарата содержат в ЭЭГ веретена.
Ряд гормональных и фармакологических веществ, снижающих тонус активирующих систем, видимо,также вызывают веретена. Особенно хорошо изучены барбитуровые веретена. Одно время полагали, что барбитуровые веретена в глубокую стадию наркоза отличаются от обычных веретен. Однако данные нашей лаборатории (Мухаметов, 1968) показали, что по мере углубления наркоза постепенно меняются и свойства веретен, что дало основание утверждать, что веретена глубокого барбитурового наркоза тоже являются модификацией ритма сонных веретен.
Спонтанные веретена регистрируются почти во всех отделах коры, таламусе и в ряде других структур мозга. Вызванные веретена, РВ и РУ могут быть приурочены к определенным ограниченным участкам мозга, что прямо связано с преимущественным возбуждением тех или иных отделов таламуса. Экспериментальный материал свидетельствует также о том, что спонтанные веретена, начинаясь в любом участке таламуса, распространяются на другие участки, а распространение активности в коре лишь отражает этот таламический процесс. Вот почему теперь, после общей характеристики разных модификаций σ-ритма. можно перейти к его генезу и прежде всего выяснять роль таламуса в его происхождении.
Таламус — водитель σ-ритма
Еще в самых ранних работах по одновременной регистрации спонтанней элекрической активности в коре и таламических ядрах было отмечено, что в них может регистрироваться однотипная и синхронная активность (Spiegel, 1937). Более поздние исследования Mopиcoнa и других (Morison, Dempsey, 1943), касающиеся веретен, относительно глубокого барбитурового наркоза показали, что синхронное, возникновение вспышек этой активности в коре и таламических ядрах (неспецифических) является характерным ее свойством. Во время естественного сна одновременное появление типичной веретенообразной активности в коре и таламусе отметили Гесс и др. (Hess et al., 1953). Обязательная приуроченность веретенообразной активности к коре и таламусу и синхронное ее появление в этих структурах дали основание для предположения, что одна из структур является первичным очагом этой активности, а в другой — веретенообразная активность имеет вторичный характер. Поскольку основные проводящие пути направлены от таламуса к коре, наиболее естественным было предположение, что веретенообразная активность первично возникает в таламусе и по таламо-кортикальным волокнам проводится в кору.
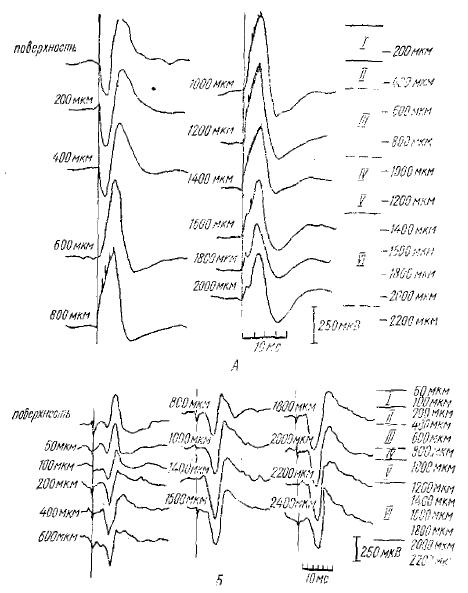
Вертикальная линия — момент раздражения релейного таламического ядра. Около каждой кривой глубина погружения электрода (мкм); I—VI — слои коры и глубина регистрации потенциалов (мкм)
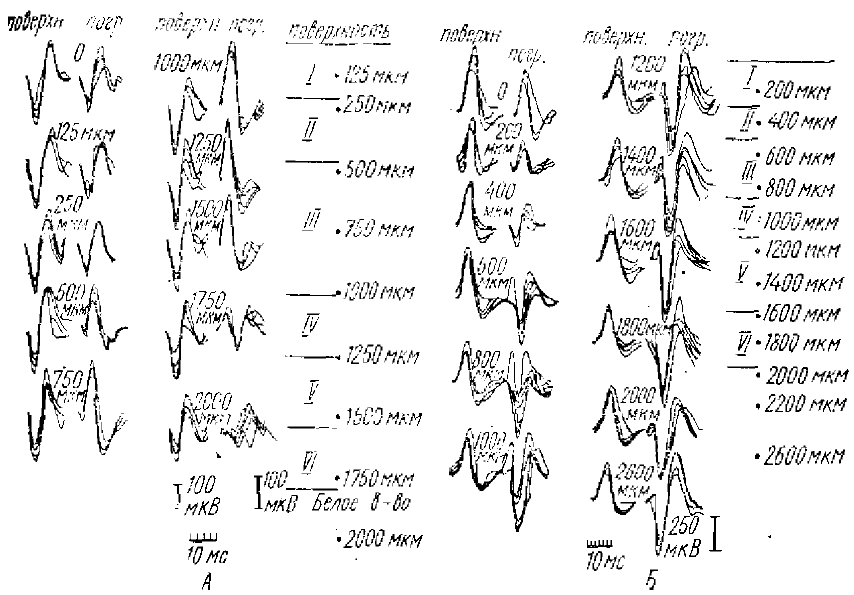
Из нескольких записей выбиралось от 4 до 6 волн данного типа, и кривые накладывались друг на друга. В каждой паре кривых справа — запись от погруженного электрода, глубина погружения указана в мкм, слева — запись от неподвижного электрода на поверхности копы, полученная одновременно с расположенной рядом глубинной записью. Остальные обозначения такие же как на рис. 2
1 — суммарные, потенциалы, регистрируемые из глубоких слоев коры, 2 — импульсная активность, 3 — суммарная активность, регистрируемая от поверхности коры
Проверка этого предположения проводилась в основном с помощью метода экстирпаций и анатомической изоляции коры и таламуса. Было показано (Morison, Basset, 1945), что после декортикации (как унилатеральной, так и билатеральной) типичные вспышки веретенообразной активности в интраламинарных ядрах таламуса могут наблюдаться до 3 дней. Исчезновение веретенообразной активности в таламических ядрах наблюдалось только через значительное время после операции (в хронических опытах до 21 дня и более), что вполне обоснованно связывается с наступлением дегенерационных процессов в таламусе. Подобные данные получили Гол и Келлуей (Gel, Cellaway, 1963; Ccllawav et al, 1966) на препарате «изолированного таламуса», где были пересечены все связи между таламусом и корой. В таком отделенном от коры таламусе можно регистрировать стойкую ритмическую активность с частотой 8—12 Гц, которая сходна с веретенообразной активностью (рис. 5). Таким образом, кора головного мозга не является необходимой для генерации ритмической активности в таламусе.

С другой стороны, как показывают опыты с разрушением или отсечением таламуса, последний является необходимым для генерации ритмической веретенообразной активности в коре, причем необходим именно сам таламус, а не другие структуры, которые могли бы задавать ритм активности через посредство таламических ядер.
Итак, самые различные хирургические вмешательства, выключающие влияние таламических ядер на кору мозга, дают один и тот же эффект — исчезновение в коре веретенообразной активности.
На основании многочисленных фактов к настоящему времени сформировалось широко распространенное мнение, что только таламус обладает способностью к автономной генерации веретенообразной активности; кора же такой способностью не обладает, и веретенообразные вспышки появляются в ней только под влиянием ритмических таламо-кортикальных залпов импульсов, генерируемых таламусом при возникновении в нем ритмического процесса. Таким образом, таламус является водителем ритмической веретенообразной активности по отношению к другим структурам. Это подтверждается и разрушением других структур мозга: хвостатого ядра, перегородки, гипоталамуса, субталамуса. Их разрушение не влияет на генерацию спонтанных и вызванных веретен (и РВ и РУ) в таламусе и коре.
Представление о первичной генерации веретенообразной активности в таламических ядрах подтверждается также и экспериментами другого рода — исследованиями влияния на частоту веретенообразной активности локальных воздействий на различные структуры мозга и прежде всего на таламус и кору.
Опыты с раздельным воздействием на кору и таламус дали вполне четкие результаты. При локальном воздействии на таламические ядра инъекции пенициллина (Ralston, Ajmon — Marsan, 1956) наблюдалось резкое уменьшение частоты веретенообразной активности, регистрируемой от коры (с 8—12 в 1 с до 3,5—5 в 1 с. на кошках под барбитуровым наркозом). Можно было наблюдать, что локализация в коре видоизмененных (замедленных) веретен зависит от локализации таламической инъекции пенициллина. Эти результаты подтверждают представления о первичном возникновении веретенообразной активности в таламусе. Локальные воздействия на кору не дают такого эффекта. По данным ряда авторов (Bremer, 1958; Andersen et al., 1967; Jasper, Shacter, 1970), изме-нение температуры поверхности коры не меняет частоты веретенообразной активности; частота снижается только при общем охлаждении тела, которое должно затрагивать и подкорковые образования. Систематическое исследование такого рода было проведено С. П. Наракашвили и сотрудниками (1964, 1965), которые исследовали изменения веретенообразной активности при воздействии в одинаковых условиях ряда веществ (как стимулирующих,так и депрессирующих) на кору и на таламические ядра. В полном соответствии с предыдущими данными было найдено, что аппликация применяемых веществ на ограниченные участки поверхности коры увеличивает или уменьшает амплитуду потенциалов веретенообразной активности, но не меняет их частоты. При инъекции растворов этих же веществ в неспецифические таламические ядра частота веретенообразной активности в коре заметно изменялась. Очевидно, что все эти результаты полностью согласуются с данными, полученными путем хирургических вмешательств, и показывают, что таламус является первичным очагом веретенообразной активности.
Однако существуют данные о наличии сильных регулирующих влияний на таламические ядра со стороны коры (Tiberlin et al., 1961 и Др.), влияющих на генерацию ритмической веретенообразной активности в таламусе. Эти данные не противоречат представлению о первичной генерации в таламусе веретенообразной активности. Наличием таких регулирующих влияний можно объяснить как прекращение генерации в таламусе ритмической активности после довольно грубого вмешательства — удаления коры, так и зависимость включения таламического генератора ритмической активности от функционального состояния коры.
В связи с этим необходимо, видимо, рассматривать и данные Веласко и Линдсли (Velasco, Lindsley, 1965), считающих, что для сохранения веретенообразной активности и реакции вовлечения необходим совершенно определенный участок коры мозга — орбитальная кора. В их опытах удаление всей дорзолатеральной коры мозга и медиальной коры не устраняло веретенообразную активность и реакцию вовлечения, но удаление одной только орбитальной коры полностью подавляло веретенообразную активность как во всех остальных областях коры, так и в таламусе. Таким образом, орбитальная кора, по мнению Веласко и Линдсли, является единственной специфичной зоной коры, играющей решающую роль в регуляции таламо-кортикальной ритмической активности.
Впоследствии многими исследователями этот вывод о роли орбитальной коры был пересмотрен. Было показано, что разные корковые поля и особенно орбитальная кора могут оказывать сильное влияние на механизм веретен, хотя сами они не являются структурами, необходимыми для их генерации. Эти влияния, видимо, могут носить облегчающий и тормозной характер на таламический механизм веретен. Что касается не спонтанных, а вызванных веретен, то анализ многочисленных литературных источников свидетельствует также о их таламическом происхождении. Это касается сенсорных послеразрядов (РРП по типу веретен), веретен хвостатого ядра и веретен на прямое раздражение коры.
Итак, суммируя все сказанное, можно прийти к заключению, что большинство экспериментальных данных указывает на первичное возникновение веретенообразной активности в таламусе; данных же, которые бы достоверно противоречили такому взгляду, пока не получено.
Теперь возникает вопрос, какие таламические ядра или группы ядер являются первичными очагами ритмической активности? Известно, что таламус содержит несколько групп ядер, имеющих различное функциональное значение — специфические, неспецифические и ассоциативные ядра. Все ли эти ядра обладают способностью к автономной генерации ритмической веретенообразной активности или эта способность является свойством только нескольких специализированных ядер или ядерных групп?
Еще Морисоном и Дэмпси было сделано заключение, что спонтанная веретенообразная активность («взрывная активность») является функцией неспецифической таламо-кортикальной проекционной системы и, следовательно; первичным очагом возникновения этой активности являются неспецифические (медиальные) таламические ядра.
Эта точка зрения получила весьма широкое распространение и подтверждение в экспериментах с разрушением таламических ядер. Было показано, что наибольшие изменения электрической активности коры вызывало разрушение неспецифических ядер таламуса — медиальных и интраламинарных; после разрушения этих ядер веретенообразная активность в коре исчезала. Меньший эффект давало разрушение передней группы ядер таламуса. Разрушение же специфических переключательных ядер таламуса вызывало минимальные изменения электрической активности коры на стороне разрушения и не устраняло веретенообразной активности. Отсутствие веретенообразной активности в коре после ограниченной коагуляции интраламинарных таламических ядер отмечали также Хёсли и Монье (Hoslv, Monniеr, 1963).
По мнению Э. С. Мониава и Р. К. Борукаева (1965), разрушение специфических релейных ядер таламуса может даже способствовать появлению веретенообразной активности в проекционной зоне коры, соответствующей разрушенному таламическому ядру. Все это безусловно указывает на основную роль неспецифических ядер таламуса (медиальной и интраламинарной групп ядер) в формировании спонтанной ритмической активности веретенообразного типа.
Однако имеется достаточно фактов, показывающих, что способностью к генерации веретенообразной активности обладают также специфические и ассоциативные ядра таламуса. Так, Кавамура и Ямамото (Kawamura, Jamomoto, 1961) показали, что в возникновении веретенообразных вспышек в слуховой коре решающую роль играет специфическое ядро слухового анализатора — медиальное коленчатое тело. Имеются и другие данные, указывающие на возможность тесной связи веретенообразной активности со специфическими ядрами таламуса. Мы уже указывали, что Спенсером и Брукхартом (1961) был сделан вывод, что в генерации спонтанной веретенообразной активности могут принимать участие как специфические, так и неспецифические таламо-кортикальные проекционные системы, т. е. эта активность может первично возникать как в специфических, так и в неспецифических таламических ядрах. В зависимости же от того, какое таламическое ядро — специфическое или неспецифическое — является ведущим в каждый данный момент, определяется соответствии со способом проекции этого ядра на корковые элементы и форма корковых потенциалов, и их принадлежность к одному из двух видов. Специфические релейные ядра таламуса, также как и неспецифические, могут быть самостоятельным первичным очагом веретенообразной активности.
Что касается относительной значимости специфических и неспецифических таламических ядер в контроле веретенообразной активности, то вполне возможно, что для большинства областей коры более значимым водителем являются неспецифические ядра, чем и объясняются изложенные выше данные, расцененные авторами как свидетельство связи веретенообразной активности с деятельностью только неспецифической таламо-кортикальной системы.
В противовес точке зрения об исключительной роли неспецифических ядер в генезе корковых веретен Андерсен и Андерссон (Andersen, Andersson, 1967, 1968) также полагают, что существует целый ряд отдельных пейсмекеров в ядрах таламуса, генерирующих автономно σ-ритм, которые не всегда вовлекаются в генерацию каждого веретена. Они нашли, что в одном таламусе, может быть около 35 000 таких генераторов, т. е. даже близко расположенные группы нейронов таламуса генерируют веретена независимо друг от друга. Тогда корковое поле, контролируемое таким пейсмекером, может соответствовать, по сути дела, размеру корковых колонок.
Явление рекрутирования волн веретен
Этот механизм вначале лучше рассмотреть на примере РВ и РУ. Здесь также существует целый ряд данных, показывающий, что именно в раздражаемом таламическом ядре происходит характерная «раскачка» ответов от стимула к стимулу при включении ритмических раздражений, благодаря которой ответы становятся по амплитуде во много раз больше, чем ответы на одиночные раздражения. Рекрутирование на таламическом уровне ведет к тому, что разряды таламических нейронов, посылающих свои аксоны в кору, становятся сильнее от стимула к стимулу и соответственно возрастает амплитуда ответов коры на эти таламо-кортикальные залпы, что и регистрируется в виде кортикальных РВ и РУ. Причем свойством «раскачки», «рекрутирования» ответов при низкочастотном ритмическом раздражении обладают как специфические (релейные), так и неспецифические ядра таламуса. Кора же головного мозга не может без участия таламуса генерировать РВ и РУ. Это не исключает активного участия коры мозга в формировании ритмических реакций типа РВ и РУ путем влияния на раздражаемое таламическое ядро. Возможность такого влияния тем более следует учитывать, что большинство работ по исследованию РВ и РУ проведено в условиях неглубокого барбитурового наркоза, который может влиять на функции коры сильнее, чем на таламические. Вполне возможно, что функциональная роль коры в формировании этих реакций выступала в разобранных работах относительно слабо, а в нормальных условиях (при отсутствии наркоза) может выявляться значительно сильнее.
Свойства и различия РВ и РУ в связи со спецификой проекции на кору мозга специфических (релейных) и неспецифических таламических ядер
Процесс рекрутирования при низкочастотных ритмических раздражения происходит, по-видимому, одинаково как в специфических, так и неспецифических ядрах таламуса. Однако залпы таламо-кортикальных импульсов, исходящие из раздражаемого ядра, могут вызвать в коре ответы совершенно различных типов в зависимости от того,.в какие зоны и слои коры приходят волокна от этого ядра. Специфические релейные ядра таламуса, как показывают морфологические и физиологические, данные, посылают волокна в III—IV слои проекционных зон коры. В соответствии с этим зона регистрации РУ ограничена только той проекционной областью коры, которая соответствует раздражаемому таламическому ядру. Форма потенциалов РУ (позитивно-негативный комплекс при регистрации от поверхности коры) качественно сходна с формой первичного ответа коры, вызываемого одиночным раздражением того же специфического таламического ядра, из которого вызывается и РУ при ритмическом раздражении. Исследование распределения по поперечнику коры потенциалов при развитии каждой волны РУ также выявило картину очень сходную с той, которая наблюдается для первичного ответа. В глубине коры регистрируются негативно-позитивные ответы, а по мере передвижения регистрирующего электрода к поверхности коры латентный период негативной фазы увеличивается и перед ней вырастает позитивная фаза, так что на поверхности ответы превращаются в позитивно-негативные (Spencer, Brookhart, 1961). Можно рассматривать потенциалы РУ как гипертрофированные первичные ответы коры, возникающие в ответ не на короткий разряд импульсов из таламического релейного ядра, возникающий при одиночном его раздражении, а на значительно усиленный и растянутый залп импульсов, который стал таким в результате внутриталамических процессов рекрутирования, активированных низкочастотным ритмическим раздражением. Но такая интерпретация отнюдь не является общепринятой. Существуют высказывания и другого порядка, что потенциалы РУ являются комбинацией двух ответов различной природы, наложенных друг на друга— первичного ответа и рекрутирующегося потенциала той же природы, что и потенциалы РВ (Bishop et al., 1961).
Корковые потенциалы РВ значительно шире распространены по коре, чем первичные ответы и потенциалы РУ, что связано с относительно диффузным характером связей между неспецифическими таламическими ядрами и корон. Правда, РВ может возникать и в относительно локальных областях коры, поскольку диффузность неспецифических таламо-кортикальных связей не является абсолютной (Jasper, 1949; Haberly, Jasper, 1953). Но по сравнению с первичными ответами и РУ реакция вовлечения является все же значительно менее локальной.
Форма потенциалов РВ также отличается от форм ответов на раздражение специфических проекционных ядер таламуса. При регистрации от поверхности коры потенциалы РВ практически монофазны (негативны). Может наблюдаться также небольшая короткая поверхностнопозитивная волна перед основным поверхностно-негативным потенциалом, а также более длительная, но низкоамплитудная вторичная поверхностно-позитивная волна после негативного потенциала, но последний можно считать основным компонентом потенциалов РВ как по признаку максимальной амплитуды, так и по признаку наибольшей стабильности регистрации. Распределение по поперечнику коры потенциалов во время развития РВ также совершенно отлично от того, что наблюдается для первичного ответа или потенциалов реакции усиления (Li et al., 1956; Spencer, Brookhart, 1961). Характерная форма корковых потенциалов РВ была расценена как указание на специфику проекции на кору волокон от неспецифических таламических ядер. Очень широко распространено мнение, что импульсы от неспецифических ядер таламуса приходят диффузно в обширные корковые области, одновременно в различные слои коры — не только более глубокие, получающие афференты также от специфических ядер, по и в поверхностные. На отличие неспецифических таламо-кортикальных проекций от специфических указывает также, разная способность РВ и РУ к горизонтальному распространению по коре. В проекционной корковой области подрезка коры, прерывающая таламо-кортикальные связи, но не затрагивающая горизонтальные внутрикортикальные связи, приводит к тому, что РУ в этом участке больше не может быть вызвана, тогда как РВ на раздражение неспецифического ядра по-прежнему регистрируется, как и в интактной коре (Hanbery, Jasper, 1953).
На В и Г — верхняя пара кривых — ответы на первый стимул, нижняя — ответы на пятый стимул
Пути передачи импульсов из неспецифических таламических ядер в кору исследовались многими авторами, так как они представляют интерес не только в связи с вопросом о механизмах РВ, по и в связи со всей проблемой организации таламо-кортикальных систем. Предполагалось, что неспецифические ядра таламуса не имеют (или почти не имеют) прямых связей с корой мозга и что проведение импульсов, вызывающих РВ в коре, происходит через ассоциативные ядра или ретикулярное ядро таламуса. Однако исследования Шлага и др. (Schlag et al., 1966) показали,, что при регистрации ответов на ритмические раздражения таламуса от орбитальной зоны коры мозга на ритмическое раздражение срединных или интраламинарных ядер таламуса (т. е. ядер, расцениваемых всеми как неспецифические) возникает ритмическая реакция, потенциалы которой по форме полностью напоминают потенциалы РУ, тогда как в остальных зонах коры при этом регистрируется обычная РВ. Наоборот, при ритмическом раздражении релейного таламического ядра в соответствующей ему проекционной зоне коры регистрировалась РУ, но в орбитальной коре в это время возникали потенциалы, которые по форме можно было бы классифицировать как РВ (рис. 6). Отсюда делается вывод о существовании локализованной проекционной системы от неспецифического таламуса к орбитальной коре.
Эти результаты поставили под сомнение правильность всей концепции существования неспецифической таламо-кортикальной системы. Если на самом деле неспецифические ядра таламуса проецируются в совершенно определенную, четко ограниченную область коры (орбитальную область) и импульсы из неспецифических таламических нейронов вызывают в этой области коры такие же реакции, какие вызывают в классических проекционных зонах импульсы из релейных ядер таламуса, то, естественно, возникает вопрос о правомерности противопоставления специфических и неспецифических таламических ядер. Такой вопрос уже неоднократно ставился в литературе и в последние годы нашел свое подтверждение в анатомических и физиологических исследованиях (Адрианов, 1964).
Таким образом, структурные основы РУ и РВ (в особенности последней) пока выяснены далеко не до конца. Довольно уверенно можно говорить о решающей роли раздражаемых таламических ядер в формировании таких наиболее характерных черт реакции, как рекрутирование ответов и фаз нарастания-спада. Но что касается путей прихода в кору залпов импульсов от таламуса и локализации в коре окончаний волокон, проводящих эти импульсы, то приходится признать, что этот вопрос разработан пока недостаточно.
Микроэлектродное изучение σ-ритма в таламусе, коре и других структурах мозга. Ценная информация о механизмах σ-ритма была получена при его микроэлектродном изучении в различных структурах мозга и прежде всего в таламусе.
При мопополярной суммарной регистрации различных спонтанных и вызванных модификаций σ-ритма практически в любом-отделе таламуса кошки имеет место сходная в основных чертах картина колебаний потенциала. Один период ритма состоит из длительной положительной волны, которой может предшествовать негативная волна меньшей амплитуды и длительности.
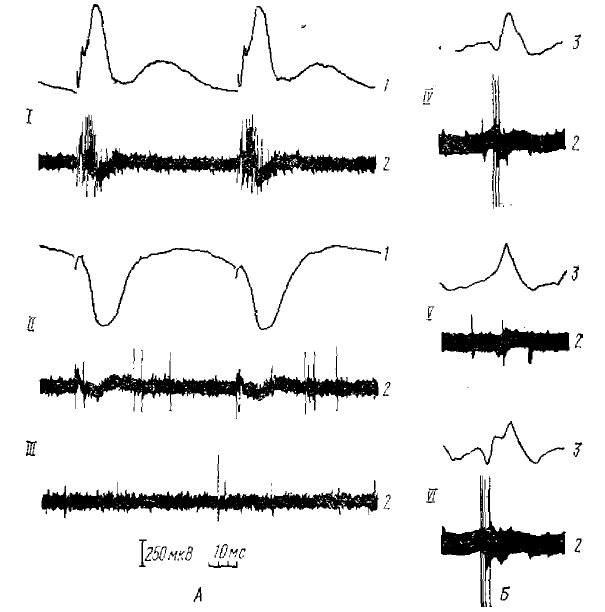
Многие таламические нейроны на фене σ-ритма начинают разряжаться ритмично и синхронно. Пачки импульсов в большей или меньшей степени приурочиваются к негативным волнам (Verzeano et al., 1954; 1958; 1960, 1961; Andersen et al., 1964, 1968, и др.). В результате внутриклеточного изучения барбитуровых веретен, специфических σ-послеразрядов (Andersen et al., 1962, 1964, 1968) и реакции вовлечения (Purpura, Cohen, 1962) было обнаружено, что положительным волнам σ-ритма соответствуют тормозные постсинаптические потенциалы (ТПСП) большинства таламических нейронов. Этим ТПСП могут предшествовать деполяризационные отклонения мембранного потенциала, сопровождающиеся импульсными разрядами; однако деполяризация и спайки имеются далеко не перед каждым ТПСП (см. рис. 9).
Мы уже говорили о последовательности синаптических процессов в ядрах таламуса (коленчатое тело, вентробазальный комплекс) после их однократной, компактной афферентной активации.
➥ Более подробно: Возбуждение и торможение в головном мозге
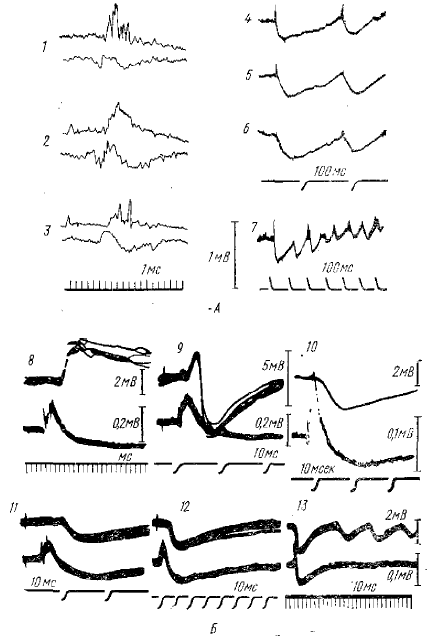
Афферентные импульсы, которые приходят по волокнам зрительного тракта и оканчиваются на переключательных клетках латерального коленчатого тела или клетках ядер вентро- базальной группы, вызывают в этих клетках ВПСП, иод действием которого переключательные клетки генерируют залп импульсов, направляющийся по таламо-кортикальным волокнам к коре головного мозга. Одновременно импульсы заходят в коллатерали таламо-кортикальных волокон, которые возвращаются к тем же переключательным клеткам и (по-видимому, через посредство вставочных тормозных нейронов типа клеток Реншоу) образуют на них тормозные синапсы. Активированные таким путем тормозные синапсы вызывают длительную гиперполяризацию (ТПСП) переключательных клеток, следствием которой является прекращение дальнейших разрядов, пока гиперполяризация не закончится и не восстановится исходное состояние нейронов.
1 — лучевого нерва, 2— срединного нерва, 3 — локтевого нерва (верхняя кривая каждой пары — первичная негативная волна ответа таламического ядра на раздражение нерва при быстрой развертке, нижняя кривая — одновременная регистрация ответа клиновидного ядра), 4, 5, 6— длинная позитивная волна и повторная вспышка импульсной активности в таламическом ядре (4 — ответ на раздражение поверхностного лучевого, 5 — срединного, 6 — локтевого нерва). 7 — ритмический разряд последействия, 8—нейрон, в котором в ответ на раздражение афферентного нерва возникает только длительный ВПСП. 9—нейрон, в котором первичный ВПСП обрывается высокоамплитудным ТПСП большой длительности, 10 —нейрон, в котором в ответ на афферентное раздражение возникает только длительный ТПСП, 11, 12, 13 — реакция нейрона, начинающаяся с длительного ТПСП, при различных скоростях развертки (в каждой паре кривых сверху — внутриклеточная регистрация активности клиновидного ядра)
Сопоставление внутриклеточно регистрируемых колебаний потенциала с суммарной электрической активностью таламических ядер (рис. 7) показывает, что в большинстве случаев описанному двухфазному (деполяризация — гиперполяризация) колебанию внутриклеточного потенциала при однократной активации таламического ядра соответствует такое же двухфазное колебание суммарного электрического потенциала ядра. Первичному ВПСП соответствует, как уже говорилось, негативное колебание суммарного потенциала. Последующий ТПСП отражается в суммарной электрической активности в виде длительной позитивной волны, которая регистрировалась авторами, обращавшими внимание на поздние компоненты электрической реакции таламического ядра на раздражение (Marshall, 1941; Vastola, 1959; Andersen et al., 1964), По форме и длительности эта позитивная волна совпадает с внутриклеточно регистрируемыми ТПСП и периодами торможения, обнаруживаемыми по угнетению спонтанной импульсной активности или по депрессии ответов на тестирующие раздражения. Так же как и внутриклеточно-регистрируемые ТПСП, эта волна может быть вызвана антидромным раздражением тала-мокортикальных волокон (Vastola, 1959; Andersen et al., 1964), в том числе и при соблюдении условий, исключающих соответствующую активацию кортико-таламических волокон. Рассмотренный комплекс из возбуждающего и тормозного постсинаптических потенциалов, который по различным признакам легко обнаруживается в таламических ядрах при одиночном афферентном раздражении, служит основой формирования также и ритмической активности нейронов таламуса.
Действительно, реакция отдельных таламических элементов на одиночные периферические раздражения не всегда ограничивается появлением одного комплекса ВПСП-ТПСП, после которого происходит восстановление внутриклеточного потенциала и импульсной активности до исходного, фонового уровня. Часто после первого комплекса ВПСП-ТПСП следуют повторные аналогичные колебания внутриклеточного потенциала, состоящие из чередующихся волн гиперполяризации (ТПСП), сопровождающихся угнетением импульсных разрядов нейронов, и волн деполяризации, сопровождающихся вспышками нейронной активности (Andersen et al., 1964), т. е. наблюдается явление, которое можно назвать внутриклеточным ритмическим разрядом последействия (рис. 7, Б, 1,3). Ритмические вспышки импульсной активности, сопровождающие эти колебания мембранного потенциала, можно регистрировать и при экстраклеточном микроэлектродном отведении.
Внутриклеточные разряды последействия, синхронно возникающие в основной массе таламических нейронов, находят непосредственное отражение в экстраклеточных суммарных потенциалах. Помимо первого негативно-позитивного комплекса, соответствующего первому комплексу ВПСП-ТПСП, в суммарной электрограмме таламического ядра могут регистрироваться повторные колебания, соответствующие повторным колебаниям клеточных потенциалов: более короткие негативные волны, являющиеся отражением внутриклеточных ВПСП, чередуются с длительными (порядка 100 мс) позитивными волнами, которые являются отражением внутриклеточных ТПСП (Andersen et al., 1964). Таким образом, регистрируемые от таламических ядер ритмические разряды последействия являются непосредственным отражением колебаний мембранного потенциала в нейронах этих ядер.
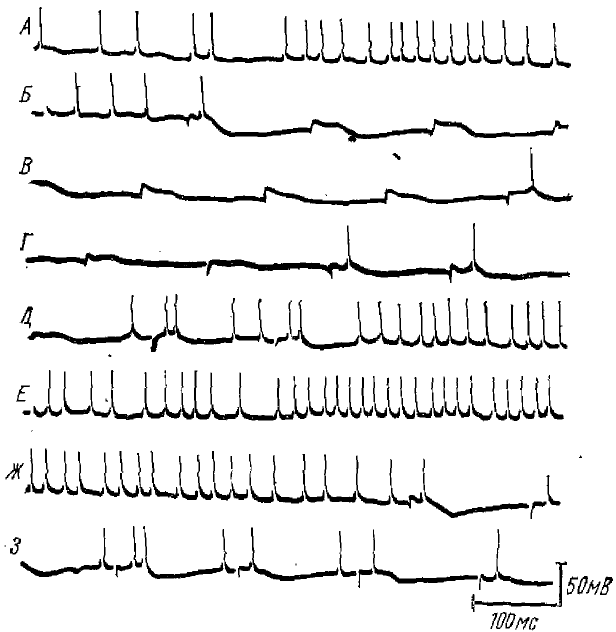
Процессы, во многом сходные с описанными выше, наблюдаются в таламических нейронах и при ритмических раздражениях ядер таламуса (рис. 8), т. е. при провокации РВ или РУ (Purpura et al., 1962, 1963).
Последовательность возбуждающих и тормозных постсинаптических потенциалов наблюдается при низкочастотном ритмическом раздражении не только в ядрах средней линии таламуса, но и в специфических релейных таламических ядрах (Purpura et al., 1965). Чередование фаз активации и торможения наблюдается также в релейных таламических ядрах при ритмических афферентных раздражениях.
Кривые — последовательные отрезки непрерывной записи. На кривых А и частично Б видна спонтанная активность. Первое раздражение с частотой 7 имп/с (Б) вызывает ВПСП и импульсный разряд, за ними следует длиннолатентный (30 мс) продолжительный ТПСП. Последующие раздражения (Б, В, Г) вызывают ВПСП и ТПСП. Прекращение раздражений (Д) сопровождается длительной деполяризацией и Повышением возбудимости (Д, Е, Ж). Возобновление ритмической стимуляции (В) с частотой 7 имп/с вновь сопровождается появлением комплексов ВПСП—ТПСП. Моменты раздражений отмечены направленными вниз артефактами
Таким образом, комплекс ВПСП-ТПСП, являющийся реакцией отдельных таламических нейронов на одиночное раздражение, можно считать также и основным элементом самых разнообразных ритмических реакций нейронов как специфических, так и неспецифических таламических ядер. Суммарные, экстраклеточно регистрируемые потенциалы таламических ядер являются непосредственным отражением этих постсинаптических потенциалов, более или менее синхронно возникающих в большом количестве нейронов (т. е. генерируются теми же синаптическими токами, которые вызывают постсинаптические потенциалы в нейронах).
Процессы в отдельных таламических нейронах во время спонтанной веретенообразной активности
Наиболее детальное исследование этих процессов проведено в работе Андерсена и Сиерса (Andersen, Sears, 1964). Ими регистрировалась (как экстраклеточно, так и внутриклеточно) активность нейронов практически во всех крупных таламических ядрах, в том числе и в ядрах средней линии таламуса (которые считаются важнейшим первичным центром спонтанной веретенообразной активности), на фоне веретенообразных вспышек, вызванных легким барбитуровым наркозом. Результаты внутриклеточных отведений от таламических нейронов показали, что в начале каждого веретена в нейронах возникает серия, четко выраженных ТПСП, нарастающих по мере развития веретена и обнаруживающих значительную временную суммацию. Длительность каждого ТПСП составляет около 120 мс, что соответствует частоте следования волн веретенообразной активности, которая в этих опытах была около 8 Гц. Импульсная активность таламических нейронов практически полностью подавлялась во время каждого ТПСП и возобновлялась на короткий период только на гребне деполяризации между двумя соседними ТПСП. Во многих приводимых осциллограммах внутриклеточных потенциалов видно (рис. 9, 10), что после каждого ТПСП наблюдается быстрый, но непродолжительный скачок деполяризации (на котором, собственно, и возникают импульсные разряды нейрона), обрываю-щийся последующим ТПСП. Форма скачка деполяризации, имеющего довольно крутой передний фронт, показывает, что едва ли он является просто восстановлением после предшествующего ТПСП; скорее всего, эта деполяризация имеет активный характер, т. е. является ВПСП.
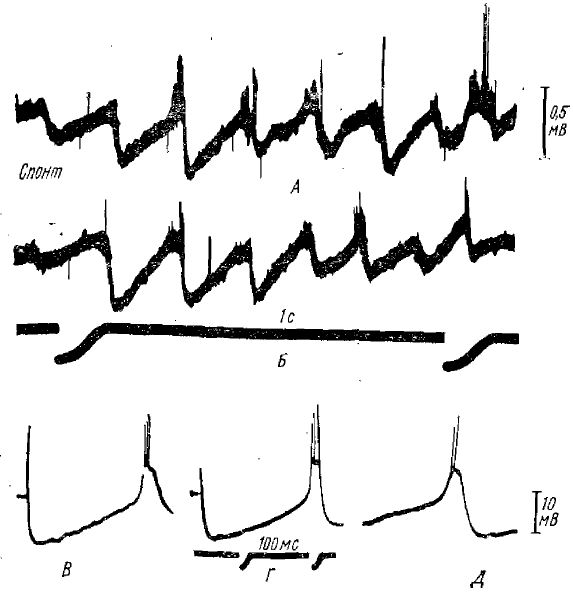
А, Б — экстраклеточные записи двух вспышек веретенообразной активности в вентральном задне-латеральном ядре таламуса; В, Г — внутриклеточные записи от нейрона вентрального задне-латерального ядра при однократной антидромной активности (раздражение белого вещества под корой); Д — внутриклеточная запись от того же нейрона во время спонтанной веретенообразной активности. Приводимый фрагмент содержит те же компоненты, что и реакция на антидромное раздражение — длительную гиперполяризацию и последующую короткую волну деполяризации с импульсными разрядами (по Андерсену и Сиерсу, 1964)
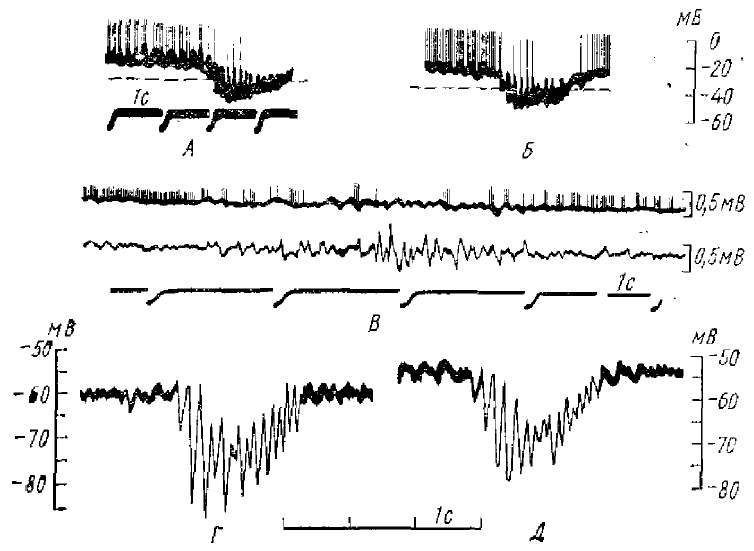
А, Б — два веретена при внутриклеточной регистрации от одного и того же нейрона вентрального задне-латерального ядра; деполяризация соответствует отклонению вверх; В—верхняя запись — экстраклеточная регистрация импульсной активности нейрона вентрального задне-латерального ядра, нижняя запись суммарная активность коры; Г, Д — внутриклеточная регистрация активности нейрона центрального латерального ядра таламуса; вспышки веретенообразной активности сопровождаются накоплением длительной гиперполяризации (по Андерсену и Сиерсу, 1964)
Следовательно, комплекс типа ВПСП-ТПСП является основным элементом внутриклеточной ритмической активности таламических нейронов не только в случае различных ритмических реакций, но и в случае спонтанных веретен.
Ритмические колебания внутриклеточного потенциала во время спонтанной веретенообразной активности происходят, по-видимому, синхронно во многих таламических нейронах, так как регистрируемые экстраклеточно суммарные электрические потенциалы находятся в хорошем соответствии с внутриклеточными потенциалами. Каждому ТПСП таламического нейрона соответствует почти такой же формы позитивная волна в экстраклеточном отведении, а пикам деполяризации между ТПСП соответствуют пики экстраклеточной негативности, на фоне которой можно зарегистрировать вспышку импульсных разрядов (Andersen, Sears, 1964). Итак, в случае спонтанной веретенообразной активности, как и при ритмических вызванных реакциях, комплексы типа ВПСП-ТПСП также играют решающую роль в формировании суммарных ритмических электрических потенциалов таламических ядер.
Аналогичные данные были получены для различных других модификаций σ-ритма: σ-послеразрядов, веретен на одиночное раздражение хвостатого ядра и коры (Andersen et al., 1962, 1964; Waszak, 1972; Гусельников и др., 1973).
Таким образом,, ритмическая активность специфических и неспецифических таламических ядер, как спонтанная, так и вызванная, сопровождается появлением в таламических нейронах чередующихся ВПСП и длительных ТПСП, причем последние имеют возвратную природу: вызываются через посредство коллатералей таламо-кортикальных аксонов к вставочных тормозных нейронов. Эти ВПСП и ТПСП, т. е. генерирующие их трансмембранные токи, находят непосредственное отражение в суммарных экстраклеточных потенциалах таламических ядер, которые обычно и регистрируются как их ритмическая электрическая активность.
Согласно гипотезе Андерсена и Экклса (Andersen, Eccles, 1962), система коллатерального торможения является основой вовлечения нейронов таламического ядра в синхронную ритмическую активность. Процесс ритмической синхронизации нервных элементов происходит, по этой гипотезе, следующим образом. Если группа нейронов таламического ядра случайно (в результате фоновой бомбардировки импульсами от других нейронов) или в результате искусственной стимуляции начнет разряжаться достаточно интенсивно и синхронно, то генерируемые ими импульсы через возвратные коллатерали аксонов возбудят вставочные тормозные клетки, послав залп импульсов к тормозным синапсам на основных (переключательных) таламических нейронах, вызовут в последних длительный ТПСП, который прервет дальнейшие импульсные разряды на время около 100 мс. Существенно, что торможение не является строго возвратным, т. е. тормозятся не только те переключательные нейроны, разрядами которых были активированы вставочные тормозные клетки, но и соседние с ними переключательные, нейроны (справедливость этого предположения подтверждается тем, что ТПСП могут возникать без предшествующих импульсных разрядов; это объясняется только существованием взаимных связей между нейронами). По окончании ТПСП вновь создается возможность появления импульсных разрядов, причем разряды в это время появляются даже с большей вероятностью, чем в фоне, до начала ТПСП. Это явление объясняется авторами гипотезы постанодальной экзальтацией — увеличением возбудимости после окончания гиперполяризации. Поскольку ТПСП возникли (благодаря наличию взаимных тормозящих связей) одновременно в значительной группе нейронов, то и заканчиваются они в этих нейронах также более или менее одновременно. Следовательно, разряды по окончании ТПСП возникают довольно синхронно в целой группе клеток, включающих как нейроны, разряды которых вызвали ТПСП, так и соседние с ними нейроны. Эти синхронно возникшие разряды приведут к повторной активаций вставочных тормозных клеток и повторному появлению ТПСП в переключательных нейронах таламического ядра. Причем, поскольку теперь повторные разряды возникают в большем количестве переключательных нейронов, чем первичный разряд, то повторная активация вставочных тормозных клеток будет более интенсивной и захватит большее количество вставочных нейронов, в результате чего повторный ТПСП также окажется более сильным, чем первый, и возникнет в еще большей группе переключательных нейронов. По окончании повторного ТПСП снова могут возникнуть разряды еще большего количества переключательных нейронов, чем после первого ТПСП, в результате чего коллатеральное торможение вызовет ТПСП как во всех тех же, так и в новых переключательных нейронах и т. д. В результате такого процесса все большее и большее количество нейронов будет вовлекаться (через посредство системы коллатерального торможения) в синхронную активность, причем эта активность будет иметь строго ритмический характер: разряд — торможение — разряд и т. д. Период ритмической активности будет, очевидно, определяться длительностью возвратных ТПСП, т. е. будет приблизительно равен 80—100 мс, что и наблюдается в действительности.
Такой механизм ритмической синхронизации активности нейронов может обеспечить как установление-спонтанной ритмической активности, так и возникновение ритмических реакций. Если система коллатерального торможения находится в достаточно активном состоянии и на ее работу не влияют внешние возмущения (что, видимо, как раз имеет место в условиях легкого барбитурового наркоза), то возможно самопроизвольное вовлечение нейронов в синхронную ритмическую активность — появление спонтанного ритма. Если синхронизация активности через систему коллатерального торможения слаба и спонтанная ритмическая синхронизация нейронов не возникает, то такая синхронизация может быть задана внешним раздражением, которое вызовет одновременный разряд значительного количества нейронов, после чего в течение некоторого времени будет поддерживаться синхронная ритмическая активность нейронов, состоящая из чередующихся вспышек активности и возвратных ТПСП, т. с. возникнет ритмический разряд последействия. Формирование реакций на ритмические раздражения таламических ядер (РВ и РУ) также легко объяснить работой системы коллатерального торможения. Каждое раздражение таламического ядра должно вызвать в его переключательных нейронах последовательность процессов изначальной активации, последующего длительного (около 100 мс) возвратного торможения и затем вторичной активации «отдачи» (постаподальной экзальтации). Если раздражать таламическое ядро ритмическими стимулами с такими интервалами, что каждый стимул будет приходиться на фазу вторичной активации после предшествующего ТПСП, то каждый последующий стимул вызовет более сильную реакцию в нейронах, чем предыдущий. Повторный стимул также может вызвать реакцию в большом количестве нейронов в результате снижения в них порога реакции после предшествующего ТПСП, вызванного коллатеральными тормозными связями от соседних нейронов. Такой процесс вовлечения,. рекрутирования нейронов в реакцию на ритмические стимулы вызовет соответствующее нарастание суммарных электрических ответов как раздражаемого таламического ядра, так и других образований (в первую очередь, коры головного мозга), в которые приходят аксоны таламических нейронов, что и наблюдается в виде хорошо известных РВ и РУ.

По абсциссе — время. по ординате — количество основных «нейронов», разряды которых зарегистрированы за 4 последовательных такта
Последующие детальные исследования активности таламических нейронов подтвердили основное значение, системы коллатерального торможения для генерации спонтанной веретенообразной активности и ритмического разряда последействия первично-таламического происхождения (Andersen et al., 1964, 1968). Моделирование на цифровой вычислительной машине (в том числе и в нашей лаборатории) нейронной сети, обладающей способностью к коллатеральному торможению, показывает, что такая система действительно может переходить в состояние спонтанной или вызванной ритмической активности, синхронной для значительного количества нейронов (Andersen, Rudjord, 1964; Шампе, Мухаметов, 1967) (рис. 11). Эти результаты являются веским подтверждением правильности основных положений гипотезы тормозной синхронизации ритмической активности таламических нейронов, разработанной Андерсеном и Экклсом.
Таким образом, гипотеза Андерсена и Экклса о происхождении ритмической активности имеет серьезное экспериментальное обоснование. Все основные исходные постулаты этой гипотезы (существование системы возвратного и взаимного коллатерального торможения; значительная длительность и величина ТПСП, вызываемых этой системой; наличие периода повышенной возбудимости по окончании ТПСП) подтверждаются физиологическими опытами и не вызывают возражений, а опыты на моделях нейронных сетей свидетельствуют о достаточности этих постулатов для возможности появления ритмической активности. Гипотеза хорошо объясняет ряд основных свойств ритмической активности таламуса (синхронизацию активности нейронов, явление рекрутирования). Наконец, объяснение механизмов установления ритмической активности отдельных нейронов на основе этой гипотезы удачно сочетается с объяснением электрогенеза суммарных относительно медленных ритмических электрических колебаний. Как было показано, эти суммарные колебания являются отражением синхронной активности тормозных синапсов, генерирующих длительные ТПСП — тех самых ТПСП возвратной природы, которые и обусловливают синхронизацию нейронной активности.
Гипотеза Андерсена и Экклса получила широкое распространение, однако в ней имеются и слабые места. Так, авторы подразумевают, что коллатеральное возвратное торможение осуществляется через вставочный нейрон. Таковыми считают обычно короткоаксонные внутриядерные таламические клетки. Однако этому нет прямых доказательств. Более того, по данным М. Е. Шейбл и А. Б. Шейбл (М. Е. Scheibel, А, В. Scheibel, 1970) в таламусе мелких грызунов, например крыс, они редки или отсутствуют. Нами же было показано (Гусельников и др., 1973), что картина мембранных процессов в таламических клетках во время веретен у крысы такая же, как и у кошки.
Таким образом, эти клетки, видимо, не являются необходимыми элементами для генерации возвратного торможения в таламусе.
Андерсен и др. (Andersen et al., 1964), Марко и др. (Marco et аl., 1967) описали клетки в таламусе кошки, по характеру активности сходные с клетками Реншоу. Однако Шлаг и Вашак (Schlag, Waszak, 1971) их не нашли. Таким образом, идентификация тормозных интернейронов еще требует своего решения.
Выдвигаются в связи с этим другие гипотезы торможения таламических нейронов, например, через ретикулярное ядро таламуса (М. Е. Scheibel, А. В. Scheibel, 1970) или путем дендро-дендритного торможения (Famigletti, 1970, 1972; Rail et al., 1966; Le Vay, 1971).
Наибольшие сомнения могут возникнуть по поводу постулата постанодальной экзальтации как причины повышения возбудимости после ТПСП в нейроне. Повышение возбудимости после периода торможения играет чрезвычайно важную роль в формировании ритмической активности, так как обеспечивает появление повторных вспышек спонтанной активности и усиление реакций на повторные раздражения, что ведет к вовлечению новых нейронов в ритмическую активность. Но надежного объяснения этому явлению пока не найдено. Механизм постанодальной экзальтации, как автоматическое повышение возбудимости нейрона после предшествующей гиперполяризации, только постулируется (Andersen et al., 1962, 1964), хотя непосредственно и не наблюдался.
Но даже если существование постанодальной экзальтации в таламических нейронах после ТПСП не подтвердится, это не будет свидетельствовать о неправильности всей гипотезы формирования ритмической активности, предложенной Андерсеном и Экклсом. Важно не существование именно постанодальной экзальтации, а важен факт обязательного повышения возбудимости нейрона после ТПСП, чем бы такое повышение возбудимости не вызывалось. Вторичное повышение возбудимости можно объяснить и другими способами. Возможно, значительную роль в образовании вторичного повышения возбудимости может играть система возвратного и взаимного возбуждения нейронов, если быстродействие ее выше, чем у системы возвратного торможения. Существование системы возвратного возбуждения, действующей несколько быстрее, чем система возвратного торможения, установлено для коры головного мозга. Для таламуса детальных данных такого рода не имеется. Но если допустить существование в таламических ядрах системы возвратного возбуждения наряду с системой возвратного торможения (такую возможность признают Андерсен и др.; Andersen et al., 1964), то повышение возбудимости нейронов таламуса после ТПСП можно объяснить точно так же, как мы объясняли аналогичное явление в нейронах коры головного мозга. А именно можно допустить, что при отсутствии синхронизированной активности таламических нейронов их фоновые разряды создают постоянный уровень активации системы возвратного торможения, которая, действуя сильнее, чем система возвратного возбуждения, ограничивает частоту импульсных разрядов на определенном уровне. Если же в нейронах синхронно возникает мощный ТПСП, то их импульсная активность на некоторое время полностью подавляется. В результате системы возвратного возбуждения и возвратного торможения могут быть активированы только после возобновления импульсных разрядов по окончании ТПСП. Но когда разряды возобновятся, то более быстродействующая система возвратного возбуждения некоторое время будет действовать без ограничения системой возвратного торможения, т. е. на короткое время включится сильная положительная обратная связь, что приведет к быстрому нарастанию деполяризации нейронов, которая может достигнуть значительной величины,— это и будет повторное повышение возбудимости нейронов после ТПСП. Такое объяснение представляется нам тем более вероятным, что на внутриклеточных записях ритмической активности таламических нейронов, приводимых в работах Андерсена, Экклса и соавторов (Andersen et al., 1964), хорошо видно резкое, нарастание скорости деполяризации в конце ТПСП, в результате чего формируется отчетливо видная волна деполяризации с довольно крутым передним фронтом, сопровождающаяся импульсными разрядами (см. рис. 9). Такое быстрое нарастание деполяризации, имеющее вид ВПСП, наиболее естественно объяснить активностью системы возвратного и взаимного возбуждения.
Возможно, существуют и другие способы объяснения вторичного повышения возбудимости таламических нейронов после ТПСП,
Можно отмстить и еще один пробел в гипотезе Андерсена и Экклса. Гипотеза очень хорошо объясняет развитие вспышки веретенообразной активности или фазы нарастания ответов РВ и РУ. Но за счет чего происходит угасание, начавшейся вспышки активности, которое в большинстве случаев неизбежно следует за фазой нарастания, а у РВ и РУ вызывает чередование увеличения и уменьшения амплитуды потенциалов?
Авторы гипотезы не приводят фактических данных, которые могли бы дать прямой ответ на этот вопрос. Однако Андерсен и Сиерс (Andersen, Sears, 1964) предполагают, что угасание вспышки ритмической активности происходит вследствие случайных флюктуаций активности отдельных нейронов, из-за которых часть нейронов начинает разряжаться не в фазе с остальными и, воздействуя на остальные нейроны, нарушает синхронность их работы.
Нарушение установившейся ритмической активности может быть следствием и других изменений, которые накапливаются именно в процессе ритмической активности и исчезают при нерегулярной активности, что дает возможность для появления новой вспышки ритмической активности (Гусельников, Супин, 1968). Таким фактором может быть накопление гиперполяризации в таламических нейронах в результате суммирования ТПСП. Действительно, внутриклеточная регистрация потенциалов таламических нейронов во время спонтанной веретенообразной активности (Andersen, Sears, 1964; Waszak, 1972; Гусельников и др., 1973) показывает, что во время «веретена» ритмические ТПСП в. нейроне возникают на фоне постепенно нарастающей гиперполяризации. Начало нарастания гиперполяризации совпадает с появлением ритмических ТПСП; это наводит на мысль, что появление- длительной гиперполяризации связано с частичным суммированием ТПСП. Нарастание длительной гиперполяризации приводит к тому, что количество импульсных разрядов в промежутках между соседними ТПСП прогрессивно уменьшается, и в конце концов разряды полностью затормаживаются. Это, естественно, сопровождается уменьшением амплитуды и исчезновением ритмических ТПСП в таламических нейронах, ритмических суммарных колебаний в таламическом ядре и ритмических ответов коры на залпы импульсов из таламуса, т. е. наблюдается фаза спада веретенобразной активности. После этого длительная гиперполяризация в таламических нейронах, не поддерживаемая более их импульсными разрядами, постепенно спадает и импульсная активность таламических нейронов восстанавливается. В начале она, очевидно, будет асинхронной и нерегулярной, так как восстановление после гиперполяризации должно происходить самостоятельно в каждом нейроне. Затем активность их может вновь синхронизироваться в результате
работы систем возвратного торможения и возвратного возбуждения — появится новое «веретено». Аналогичные процессы могут происходить и в процессе ритмической стимуляции таламических ядер, обусловливая появление фаз нарастания-спада реакций вовлечения и усиления.
Разумеется, предлагаемое нами (Гусельников, Супин, 1968) объяснение механизма спада развившейся ритмической активности является в значительной степени гипотетическим. По-видимому, возможны и другие объяснения, правильность которых могут установить только специальные эксперименты.
Генез корковых веретен σ-ритма
Многочисленные работы относительно связи веретен спонтанного σ-ритма и ее искусственно вызываемых аналогов (РВ и РУ) дают возможность построить довольно полную картину импульсных и синаптических процессов в корковых нейронах, являющихся основой этих видов активности.
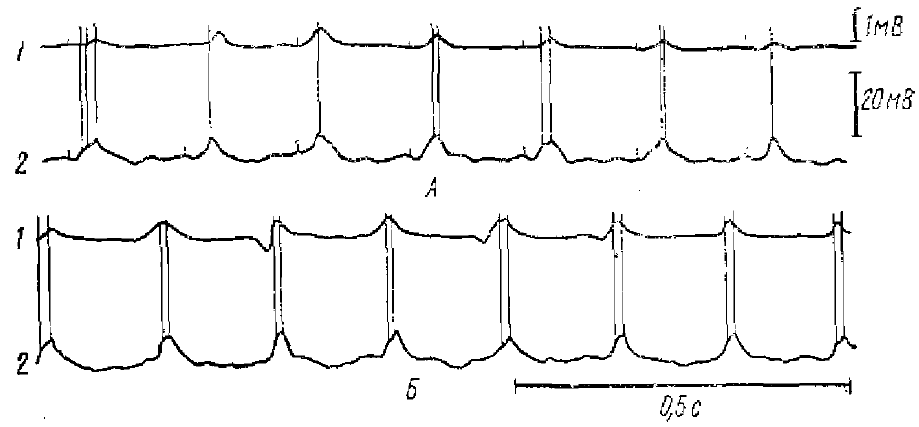
1 — суммарная активность; 2 — внутриклеточная активность
Естественно, что вначале следует рассмотреть механизмы формирования компонентов РВ и РУ, а затем спонтанных веретен σ-ритма. Наиболее известная форма потенциалов РВ, регистрируемых от поверхности коры,— негативные волны, возникающие в ответ на каждый стимул (рис. 12). Эти волны, очевидно, являются отражением активности возбуждающих (деполяризующих) синапсов, активируемых импульсами, приходящими от неспецифических ядер таламуса. Экстраклеточная негативность, регистрируемая от поверхности коры, указывает на поверхностное расположение возбуждающих синапсов, активируемых при РВ. Принято считать, что при раздражении неспецифических ядер таламуса импульсы приходят к обширным разветвлениям апикальных дендритов пирамидных нейронов коры: деполяризация и усиление импульсной активности на каждой волне РВ наблюдается не только для поверхностно расположенных нейронов, но и для нейронов, тела которых находятся в глубоких слоях коры. С таким представлением согласуются также, данные об изменении синаптических потенциалов корковых нейронов при искусственном изменении внутриклеточного потенциала: ВПСП, возникающие в ответ на раздражение срединного таламуса, изменяются при поляризации тела нейрона через внутриклеточный микроэлектрод значительно меньше, чем ВПСП, вызываемые раздражением специфического (вентро-латерального) ядра. (Creutzfeldt, Lux, 1964). Такое различное поведение, специфических и неспецифических ВПСП указывает на разное положение генерирующих их синапсов. По-видимому, неспецифические ВПСП генерируются синапсами, отделенными от тела нейрона значительным электрическим сопротивлением, что как раз и характерно для таких дендритных разветвлений.
Менее ясна природа позитивного колебания, которое может наблюдаться перед основной негативной волной РВ. Совпадение его во времени с ВПСП нейронов коры (Li, 1963) даст основание предположить, что оно является дипольным отражением активности деполяризующих синапсов, расположенных в глубине коры (направленный внутрь клетки ток вызовет в этом случае, негативный сдвиг экстраклеточного потенциала глубоких слоев коры, по отношению к которым поверхностные слои приобретут положительный потенциал). Но такое предположение пока ничем не доказано.
Неясна также природа и связь с суммарной электрической активностью ТПСП, которые могут наблюдаться в корковых нейронах при РВ. Генерируются ли эти ТПСП синапсами, образованными на нейронах коры афферентными волокнами, или они, подобно длительным ТПСП в таламусе, являются следствием работы системы возвратного торможения?
С. П. Нарикашвили и др. (1965) нашли, что импульсная активность корковых нейронов в фазу нарастания РВ в среднем увеличивается по сравнению с фоном, а в фазу спада тормозится. Этот факт расценивается авторами как свидетельство существования тормозящих таламо-кортикальных связей, деятельность которых проявляется в фазу спада РВ. Кроме того, наблюдаемое угнетение активности нейронов может быть и следствием временной суммации возвратного торможения в коре. Этот вопрос требует дальнейшего уточнения. Не установлено также четкого соответствия между внутриклеточными ТПСП и каким-либо определенным компонентом суммарных потенциалов РВ.
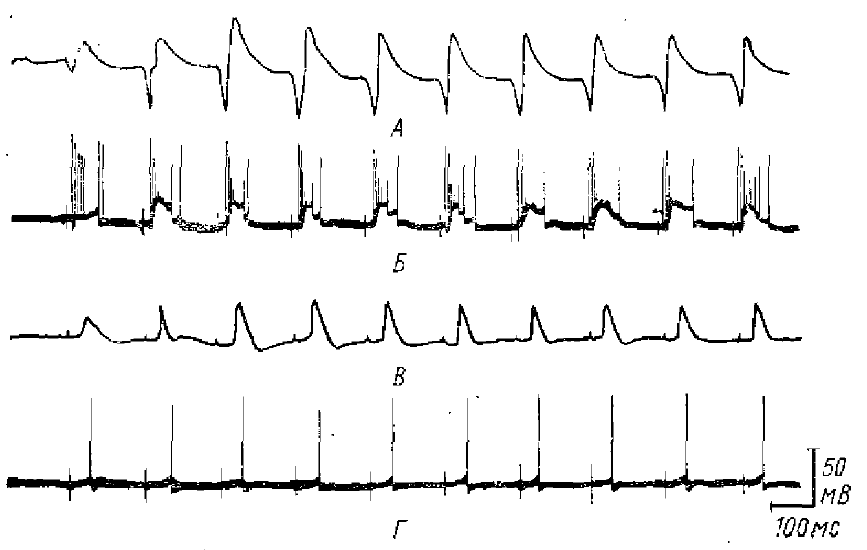
А — суммарная активность поверхности коры при ритмическом (7 имп/с) раздражении специфического таламического ядра, Б — сопровождающие эту активность (РУ) внутриклеточные потенциалы (пирамидного нейрона); В, Г — то же, что А и Б, но при раздражении неспецифических таламических ядер, т. е. при возникновении РВ (по Пурпура и др., 1964)
Кортикальные потенциалы РУ, возникающие при ритмическом раздражении переключательных специфических ядер таламуса, также сопровождаются характерными колебаниями внутриклеточных потенциалов (рис. 13).
Верхняя линия — отметка включения гиперполяризующего тока; средняя — регистрация от поверхности коры во время РУ; нижняя—внутриклеточная запись активности коркового нейрона. Искусственно вызванная гиперполяризация нейрона приводит к изменению полярности вызванного колебания клеточного потенциала; гиперполяризация (ТПСП) замещается деполяризацией.
По данным Ли (Li, 1963), основным компонентом внутриклеточной реакции на ритмическое раздражение вентрального задне-латерального ядра таламуса являются, как и при РВ волны деполяризации, ВПСП, сопровождающиеся вспышками импульсной активности нейрона. Отличие от ВПСП при РВ заключается в значительно более коротком латентном периоде. РУ может сопровождаться и внутриклеточными ТПСП (Purpura, Shofer, 1964) (рис. 14). Однако амплитуда их невелика. Даже если реакция на одиночное раздражение специфического таламического ядра содержит ТПСП значительной величины, то при переходе от одиночных раздражений к низкочастотным раздражениям наблюдается уменьшение ТПСП (Purpura et al., 1964), тогда как ВПСП нарастают от стимула к стимулу.
Отличие РУ на ритмическое раздражение от первичного ответа на одиночное раздражение специфического таламического ядра заключается, по-видимому, не только в количественном изменении величины постсинаптических потенциалов. Кли и Оффенлох (Klee, Offenloch, 1964), исследуя изменения внутриклеточных реакций при переходе от одиночных раздражений специфического таламического ядра к ритмическим раздражениям, показали, что образование реакции сопровождается появлением добавочного ВПСП, который не наблюдается при одиночных раздражениях. ВПСП, наблюдаемый при одиночных стимулах, также не остается неизменным, — при переходе к ритмической стимуляции он заметно увеличивается, что сопровождается усилением вызываемой им импульсной активности. Но помимо этого первого ВПСП ритмические раздражения вызывают второй ВПСП, следующий непосредственно вслед за первым и также сопровождающийся импульсными разрядами. Увеличение амплитуды второго ВПСП по мере ритмической стимуляции сопровождалось соответствующим увеличением отрицательной фазы суммарных потенциалов РУ регистрируемых от поверхности коры. В соответствии с наличием двух раздельных ВПСП наблюдается два максимума импульсной активности нейронов коры во время каждой волны РУ — приблизительно на 4-й и 16-й мс после стимула. Эти данные позволили Кли и Оффенлоху прийти к заключению о двойственной природе корковых потенциалов РУ. Поверхностно-позитивная фаза РУ является, очевидно, аналогом первичного ответа на одиночное раздражение, а поверхностно-негативная — ответом другого типа, сходным с поверхностно-негативными волнами РВ.
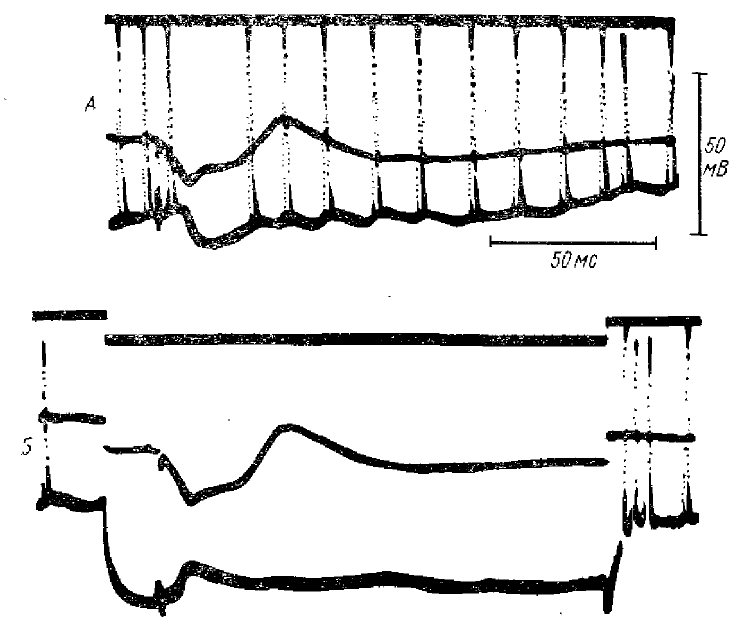
Рис. 14. ТПСП в нейронах коры, сопровождающие РУ и выявление этих ТПСП путем искусственной гиперполяризации нейрона
А — суммарный и внутриклеточный ответы, вызываемые стимулом из серии ритмических раздражений вентро-латерального ядра таламуса с частотой 6 имп/с (во внутриклеточной записи наблюдается гиперполяризационное колебание, сопровождающееся блокадой импульсных разрядов); Б — такое же раздражение на фоне искусственной гиперполяризации нейрона током 0,2 мка (по Пурпура и Шофер, 1964)
Обе фазы суммарных потенциалов РУ являются, как показывают результаты внутриклеточной регистрации, отражением деятельности возбуждающих синапсов, но локализация синапсов, генерирующих поверхностно-позитивную и поверхностно-негативную фазу, должна быть, если принять интерпретацию Кли и Оффенлоха, различной. Поверхностно-позитивные потенциалы коры могут вызываться деполяризующими синапсами, если они расположены в глубине коры, на глубинных частях вертикально ориентированных нейронов. Поверхностнонегативные потенциалы могут вызываться поверхностно расположенными деполяризующими синапсами, поскольку генерируемая ими деполяризация регистрируется в глубоко расположенных частях нейронов. Можно допустить, что эти синапсы локализованы на поверхностных разветвлениях апикальных дендритов пирамидных нейронов коры, так же как и синапсы, генерирующие поверхностно-негативные волны РВ.
Связь активности одиночных элементов коры с суммарной спонтанной ритмической активностью веретенообразного типа
В работах ряда авторов отмечается появление ритмических вспышек импульсной активности нейронов, частота которых соответствует частоте, регистрируемой в коре спонтанной веретенообразной активности. Правда, многие авторы отмечают, что соответствие вспышек импульсной активности отдельными волнами веретена довольно слабое. Как правило, импульсные разряды нейронов коры приурочены в основном к поверхностно-негативным волнам спонтанной веретенообразной активности.
Детальное исследование внутриклеточных колебаний потенциала в связи с суммарными корковыми потенциалами спонтанной веретенообразной активности было проведено на нейронах пирамидного тракта кошки Джаспером и Стефанисом (Jasper, Stefanis, 1965). Веретенообразная активность, регистрируемая от поверхности коры, состояла преимущественно из негативных колебаний. Одновременная регистрация поверхностной ЭКОГ и внутриклеточных потенциалов показала, что между ними наблюдается соответствие. Обычно с каждой поверхностно-негативной волной спонтанной веретенообразной
активности совпадала волна деполяризации в корковых нейронах. На вершине волны деполяризации возникали импульсные разряды (см. рис. 12).
Эти деполяризационные волны являются наиболее характерным компонентом внутриклеточной веретенообразной активности; гиперполяризационные колебания, хотя и могут наблюдаться, но связаны с волнами веретен менее четко.
Отсюда можно заключить, что основную роль в генерации корковых потенциалов спонтанной веретенообразной активности играют деполяризующие синапсы, расположенные в поверхностных слоях коры. Как уже указывалось ранее, картина распределения потенциалов спонтанных веретен по поперечнику коры вполне сходна с картинами, наблюдаемыми либо для волн реакции вовлечения, либо для волн реакции усиления (Spencer, Brookhart, 1961), Внутриклеточные же колебания потенциала во время волн веретенообразных вспышек также совпадают с внутриклеточными потенциалами, характерными для РВ и РУ. Следовательно, все выводы, касающиеся механизмов генерации суммарных потенциалов РВ и РУ, могут быть приложены и к потенциалам спонтанной активности веретенообразного типа. Видимо, ритмические залпы импульсов, возникающие в первичном очаге веретенообразной активности — таламусе, приходят к поверхностным слоям различных зон коры мозга, где они активируют деполяризующие синапсы, локализованные на разветвлениях апикальных дендритов пирамидных корковых нейронов, следствием чего является возникновение волны деполяризации (ВПСП) в нейронах и негативного сдвига экстраклеточного потенциала в верхних слоях коры. Этот тип генерации потенциалов веретенообразной активности, по-видимому, имеет место в большинстве случаев и, как правило, создает основной рисунок веретенообразной активности коры. Наряду с таким источником потенциалов веретен, безусловно существуют и другие источники. Это прежде всего относится к волнам веретен, которые имеют распределение потенциалов по поперечнику коры, сходные с таковым потенциалом РУ. Характер волн может быть обусловлен участием деполяризующих синапсов, локализованных в глубоких слоях коры. Наконец, не исключено участие в генерации спонтанных веретен и тормозных синапсов. Таким образом, имеется несколько различных синаптических систем, принимающих участие в генерации веретенообразной активности.
Такой вывод подтверждается детальными исследованиями распределения по поперечнику коры потенциалов веретенообразной активности (Calvet, Scherrer, 1961; Calvet, 1961; Calvet et al., 1964).
Значительно хуже, чем в таламусе и коре, изучены синаптические механизмы веретен в других нервных структурах.
В нашей лаборатории этому вопросу было уделено специальное внимание. В работе Л. М. Мухаметова и Хори (1967) было показано, что у одиночных нейронов хвостатого ядра крысы импульсные разряды были четко приурочены к определенным фазам волн веретен барбитурового сна. Дальнейшее исследование (Изнак, 1971; Gussclnikov et al., 1973) этого вопроса с использованием экстра- и внутриклеточной регистрации нейронов хвостатого ядра крысы показало, что в основе веретен лежат ритмические ВПСП многих клеток. Гиперполяризация между ритмическими ВПСП очень слаба. В отличие от таламуса кошки и крысы нейроны хвостатого ядра дают разряды на позитивных пиках волн барбитуровых веретен и веретен хвостатого ядра. В гипоталамических ядрах крысы тоже были обнаружены нейроны (Гончарская, Изнак, 1970), коррелирующие свои разряды с волнами барбитуровых веретен. Что касается других структур, то вопрос о корреляции нейронной активности с веретенами неясен, а отсюда неясен и нх генез. Так, Ружель и др. (Rougeul et al., 1966), Бальцано и Жанеро (Balzano, Jeannerod, 1970) нашли значительное увеличение импульсных разрядов во время сонных веретен в ретикулярной формации продолговатого мозга и ядрах шва, однако фазовой приуроченности разрядов нейронов во время веретен для ряда подкорковых структур не найдено (Junge, Sveen, 1968).
Итак, мы рассмотрели механизмы σ-ритма (или веретенообразной активности мозга). Мы убедились в том, что σ-ритм может быть запущен (как и другие ритмы ЭЭГ) электрическим раздражением мозга. Его можно получить раздражением самых различных отделов мозга, которое включает, видимо, в любом случае таламическую систему, генерирующую его. Иными словами, механизм запуска σ-ритма может быть сведен к искусственному
Созданию электрическим стимулом синхронности синаптических процессов в таламических нейронах — запуску ритмически чередующихся фаз возбуждения и торможения. А поскольку в этом случае эффективны и ортодромное и антидромное и прямое раздражение нейронов таламуса, понятна и широкая возможность запуска механизма σ-ритма при раздражении столь обширных участков головного мозга, с которыми таламус имеет самые тесные связи.
В настоящее время функциональная роль механизма σ-ритма остается неизвестной. Можно высказать лишь некоторые предположения. Большая часть афферентной информации, поступающей в кору, проходит через таламические ядра. Поэтому существенная перестройка нейронной активности в таламусе при возникновении сонных веретен не может не влиять на афферентацию коры. Хорошо известно, что свойства внешних раздражителей кодируются на уровне рецепторов прежде всего частотой импульсации и пространственным распределением возбуждения. Эти же свойства являются, видимо, важными характеристиками нервного сигнала. Однако для ряда видов стимуляции частотная и пространственная характеристики афферентного потока к коре должна существенно разрушаться на уровне таламических ядер во время сонных веретен, которым соответствуют синхронные изменения возбудимости таламических нейронов с четкими тормозными паузами. Следовательно, во время σ-веретен должно иметь место снижение информационного содержания афферентного потока к коре (Гусельников, Мухаметов, 1969). Кроме того, на фазе убывания (waning) σ-вспышек типичным для таламических клеток является ослабление фаз облегчения между ТПСП, что свидетельствует также о притормаживании проведения через таламус. Поэтому можно рассматривать сонное веретено как частичную функциональную деафферентацию коры. Интересно сопоставить это обстоятельство с блокадой создания, с которой коррелирует появление сонных веретен. Так, Демент и Клейтман (Dement, Klеitman, 1957) считают, что у здоровых людей в естественных условиях полное выключение сознания наблюдается только в стадиях сна, характеризующихся сонными веретенами.
В генерацию σ-ритма вовлекается целый ряд структур пирамидной и экстрапирамидной нисходящих систем.
При появлении веретен резко изменяется деятельность нейронов в этих образованиях, что заставляет предполагать существенное влияние механизма σ-ритма на эффекторные функции мозга. Однако такие нисходящие воздействия на спинномозговые и стволовые эффекторные центры практически не исследованы. Только единичные работы (Bremer, 1941; Hongo et al., 1963, и др.) связаны с этим вопросом, который, несомненно, заслуживает самого серьезного внимания.
Перспективным направлением в выяснении функционального значения σ-ритма является изучение роли его механизма в развитии сна под влиянием низкочастотного электрического раздражения мозга (Muchametov, 1974). Можно предполагать, что засыпание, наблюдающееся при стимуляции мозга по Гессу, обусловлено функционированием механизма σ-верётен, который активируется такой стимуляцией. Данное предположение является гипотезой и не может быть сейчас доказано, однако и его пользу говорят следующие данные. Для развития сна оптимальны те частоты раздражения диэнцефалических структур, которые специфичны для возникновения вызванного σ-ритма. Наилучший гипногенный эффект получается при раздражении в области неспецифического таламуса (Hess, 1944). С другой стороны, раздражение именно неспецифического таламуса даст наиболее широко распространенную вызванную модификацию σ-ритма — РВ. Поведенческое засыпание может быть вызвано раздражением многих структур (Roclofs et al., 1963; Parmeggianj, 1964, и др.). Как следует из обширного электрофизиологического материала σ-ритм тоже может возникать при электрическом раздражении большинства этих структур. По данным некоторых авторов, вызов РВ у бодрствующих животных (Jamaguchi et al., 1964; Ройтбак, Эристави, 1965) или у людей (Ischikawa, 1957) сопровождается развитием сна. Особенно важна работа Бухвальда с соавторами (Buchwald et al., 1961), которые при раздражении хвостатого ядра бодрствующих кошек получили совпадение порогов поведенческого торможения и индукции σ-послеразрядов. Все это делает высказанное выше предположение заслуживающим экспериментальной проверки
В то же время следует иметь в виду, что фактический материал, относящийся к данному вопросу, весьма сложен и крайне противоречив. Например, некоторые авторы не обнаружили поведенческого засыпания при индукции реакции вовлечения у бодрствующих кошек (Evarts, Magoun, 1957) или вызывали засыпание раздражением, не сопровождающимся потенциалами РВ (Akimoto, 1956). Кроме того, ряд существенных вопросов не изучен. Если в результате исследований окажется, что засыпание, по Гессу, действительно обусловлено активацией механизма σ-ритма, то это позволит понять многое в явлениях засыпания и сна и откроет путь к выяснению функциональной роли σ-ритма, а также можно будет предположить, что его механизм активно поддерживает и углубляет сонное состояние мозга (Гусельников, Мyxaметов, 1969).