Каков бы ни был источник суммарной электрической активности головного мозга, безусловно он должен быть тесно связан с процессами возбуждения и торможения в ЦНС, имеющими место в одиночных нервных элементах. По этому рационально вначале кратко напомнить некоторые механизмы возбуждения и торможения нервных клеток.
Из всех трех составных частей нейрона: тела клетки, дендритов и аксона, последний, являющийся наиболее простым объектом для исследования, был подробно и раньше всех изучен. Почти до 40х годов прошлого столетия большинство исследователей полагало, что закономерности деятельности аксона могут быть перенесены на другие нервные возбудимые образования и быть использованы в объяснении электрических колебаний мозга, описанных в то время.
К характерным свойствам потенциала действия (ПД) аксона относятся: возникновение его при электрической (а также химической и механической) стимуляции нервного волокна, бездекрементное проведение по нервным волокнам на большие расстояния, наличие абсолютного рефрактерного периода во время ПД и развитие его по закону «все или ничего». Постоянными значениями амплитуды и продолжительности процесса возбуждения обусловливается дискретность нервного импульса.
Для развития процесса возбуждения нервного волокна необходимым условием является поляризованное состояние мембраны, которое происходит за счет неравномерного распределения заряженных ионов по обе ее стороны; причем наружная поверхность мембраны электроположительна по отношению к внутренней. Поляризация мембраны выражается в виде мембранного потенциала или потенциала покоя.
В 1902 г. Бернштейн впервые высказал предположение, что причиной возникновения такого заряда является неравномерное распределение ионов натрия и калия. Впоследствии было точно установлено, что в состоянии покоя калий концентрируется внутри волокна, а натрий— снаружи. Так как эти ионы, в отличие от органических ионов, потенциально способны проникать в обоих направлениях через клеточные мембраны, с ними, в первую очередь, стали связывать генерацию ПД (или спайка).
Ионная природа ПД была доказана Ходжкином, Хаксли и другими исследователями. При определенном уровне деполяризации мембраны, вызванном тем или иным способом, наступает повышенная ее проницаемость к ионам натрия. Так как ионы натрия начинают двигаться в сторону их электрохимического градиента, т. е. внутрь волокна, это еще более усиливает деполяризацию мембраны и соответственно еще больше повышает ее проницаемость для ионов натрия.
Этот процесс, происходящий примерно в течение 1 мс и имеющий лавинообразный «взрывной» характер, изменяет поляризацию мембраны, уничтожая внутреннюю негативность, и даже создает небольшую позитивность на внутренней стороне мембраны.
Поступлением ионов натрия в волокно объясняется восходящая фаза ПД. С входящим натриевым потоком возникает выходящий калиевый поток. Выходящий поток калия развивается во времени в 60 раз медленнее, чем входящий натриевый поток. Подъем его происходит с постепенным нарастанием, а спуск — по экспоненте. Постоянная времени — 5-10 мс. На гребне пика начинают проявляться медленные изменения, возникающие в результате деполяризации; натриевая проводимость уменьшается, а калиевая растет. Количество ионов калия, покидающих волокно, превосходит количество ионов натрия, входящих в него; возникает процесс «инактивации», проницаемость к натрию исчезает. Это вызывает возвращение мембранного потенциала к потенциалу равновесия калия. Рост калиевой проводимости и инактивации переносящей системы натрия удерживается на несколько мс после пика и составляет основу рефрактерного периода мембраны.
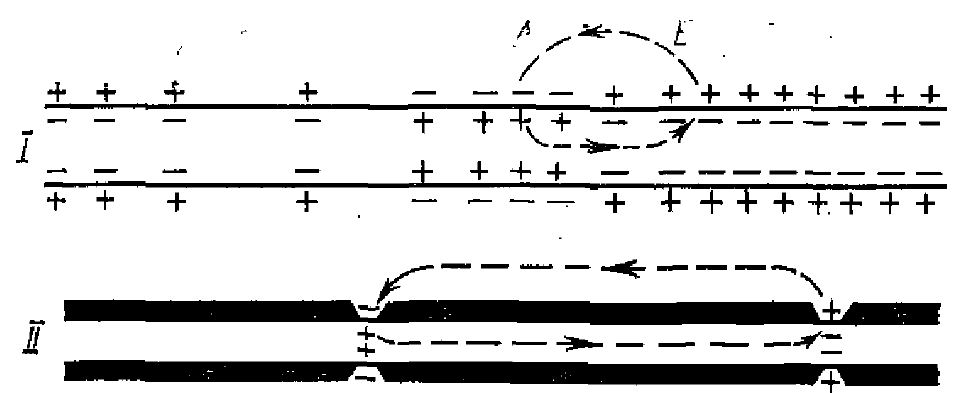
Предполагается, что причиной самораспространения нервного импульса вдоль волокна являются местные вихревые электрические токи, возникающие на границе мембраны впереди импульса (рис. 1). На рис. 1 точка А в немиэлинизированном волокне является активной, а точка Б — покоящейся. Точка А проницаема к натрию, по этому внутренняя мембрана положительна; точка Б проницаема к калию — внутренняя мембрана отрицательна. Отсюда ток течет по контуру между точкой Б и активной точкой А. Внутри аксона ток течет по направлению импульса, а снаружи против движения. Этот вихревой, местный ток в передней части активной области уменьшает заряды на емкости мембраны и тем самым уменьшает мембранный потенциал. Если разница уменьшится примерно на 20 мВ, резко увеличится и проницаемость к натрию и точка Б станет активной и может стимулировать следующую область. В миэлинизированных волокнах ток течет, видимо, от перехвата к перехвату (рис. 1, II), в результате него ПД аксона как бы «прыгает» через 2 мм, что ускоряет его распространение. Необходимо отметить, что амплитуда ПД в 5—7 раз превышает пороговую величину для активации соседнего перехвата Ранвье. Это соотношение получило название «гарантийного фактора» генерации нервного импульса (Тасаки, 1957).
По мере накопления данных по изучению возбудимых образований оказалось, что наиболее общими, свойственными всем возбудимым образованиям ответами являются градуальные потенциалы. Эти потенциалы были названы так потому, что их интенсивность меняется в зависимости от интенсивности раздражения. Кроме того, они являются локальными ответами, проводятся с декрементом и не характеризуются абсолютной рефрактерностью.
Такие потенциалы могут суммироваться. Прежде всего это касается потенциалов, регистрируемых во всех постсинаптических возбудимых образованиях при ортодромном раздражении.
Поскольку эти локальные потенциалы вызываются только под действием импульсов, при ходящих к синапсам, и генерируются в участках постсинаптической мембраны нейрона, они были названы постсинаптическими потенциалами (ПСП). В одних клетках под воздействием транссинаптического возбуждения возникает лишь постсинаптический градуальный потенциал.
В других — постсинаптический потенциал, достигнув определенного критического уровня, вызывает возникновение в клетке бездекрементного пикового потенциала (потенциала действия клетки). Электрогенез этих двух видов потенциалов различен, так как в их основе лежат различные ионные механизмы.
В ответ на выделение медиатора из пресинаптического окончания, субсинаптическая мембрана (участок мембраны нейрона, непосредственно прилегающий к пресинаптическому образованию) изменяет свою проницаемость по отношению к ионам, входящим в состав внутри клеточной и внеклеточной среды. Изменения проницаемости субсинаптической мембраны и ионные механизмы имеют неодинаковый характер при возбуждении и торможении в мозге. Возбуждение в нервной клетке связано с деполяризацией мембраны. Медиатор, освобождаемый пресинаптическим окончанием, диффундирует через синаптическую щель и вступает в реакцию с хеморецептором субсинаптической мембраны. В результате этой реакции возникает локальный процесс деполяризации. Этот деполяризационный ответ нейрона на возбуждающую стимуляцию и получил название возбуждающего постсинаптического потенциала (ВПСП). Впервые внутриклеточный ВПСП в нейронах млекопитающих был записан Броком и др. (Brock et al., 1952).
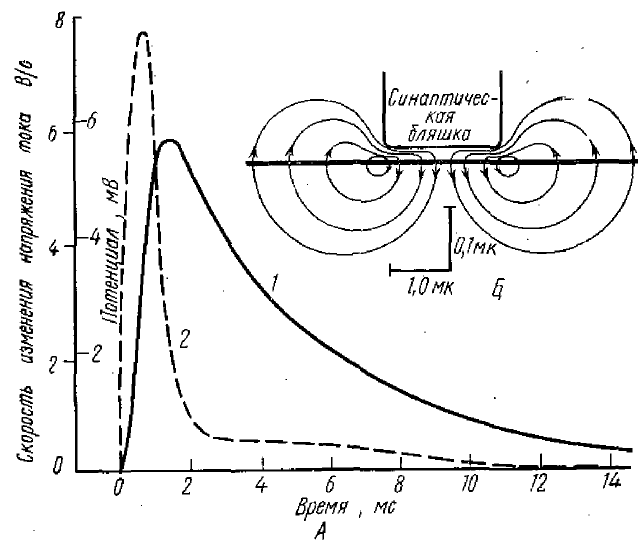
Поскольку изменения проницаемости под действием медиатора возникают в субсинаптическом участке мембраны нейрона, направление возникающего в этих частях мембраны тока будет входящим (рис. 2, Б). При этом, естественно, суммарное количество положительных зарядов внутри клетки будет возрастать. В остальных отделах мембраны (внесинаптических, или по обычной терминологии в постсинаптических) ток будет иметь выходящее направление и при его прохождении через постсинаптическую мембрану (которая обладает высоким сопротивлением порядка 1 МОм) внутри клетки возникает положительный потенциал, регистрируемый как ВПСП (рис. 2, 1). Кумбс и др. (Coombs et al., 1965) рассчитали, что создающий ВПСП ток круто возрастает до максимума в течение 0,5 мс и затем быстро убывает в течение примерно 1 мс, составляя спустя 2 мс около 10% максимальной величины (рис. 2, 2).
Величина ВПСП зависит от уровня мембранного потенциала. Если искусственно, с помощью двухканального микроэлектрода пропускать деполяризующий ток через один из его каналов, а через второй регистрировать изменения мембранного потенциала, то ВПСП изменяется прямо пропорционально уровню мембранного потенциала.
Интимный механизм реакции между медиатором и хеморецепторами клеточной мембраны до сих пор до конца не выяснен, однако реакция протекает очень быстро, а выделившийся медиатор быстро разрушается специфическими энзимами или диффундирует через синаптическую щель в окружающую среду. Не все известно и о ионных механизмах ВПСП. Возбуждающий медиатор должен резко повышать ионную проводимость субсинаптической мембраны, а единственными видами ионов, которые могут участвовать в проведении, являются ионы натрия, калия и хлора. Фэтт, Кац (Fatt, Katz, 1951) и другие исследователи в работе на концевой пластинке нервномышечного препарата предположили, что медиатор (в данном случае ацетилхолин) вызывает резкое повышение проницаемости субсинаптической мембраны, вследствие чего происходит «закорачивание» мембраны,— явление, аналогичное короткому замыканию или временному короткому проколу мембраны.
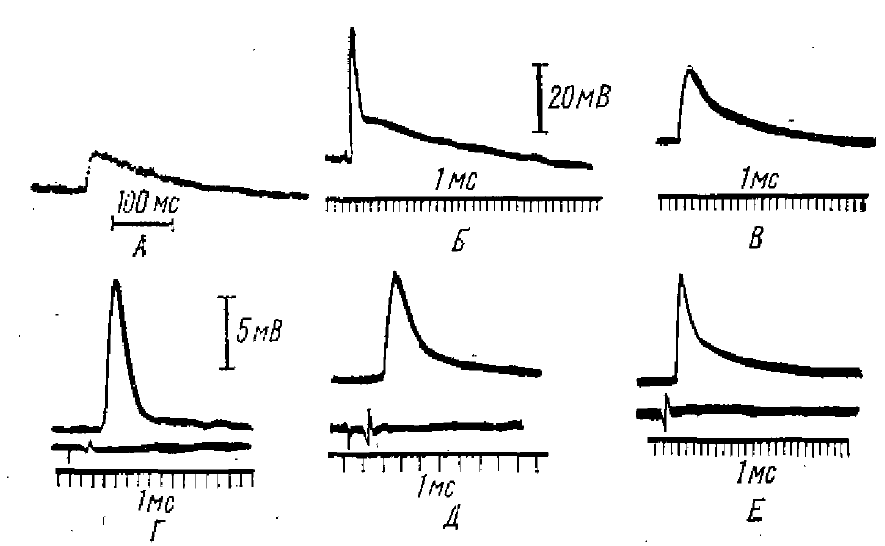
Экклс (1966) указывал, что время до развития максимума ВПСП служит мерой длительности интенсивной фазы действия медиатора на субсинаптическую мембрану. Длительность же спада ВПСП в большей степени определяется электрической постоянной времени мембраны, но может увеличиваться и за счет остаточного действия медиатора. На рис. 3 приведены примеры некоторых ВПСП возбудимых образований. Время до достижения вершин варьирует от 1 до 20 мс, а постоянная времени спада от 1 до 120 мс. К наиболее быстро «работающим» принадлежат нейроны центральной нервной системы млекопитающих. Однако и у млекопитающих синаптическое возбуждение корковых пирамидных нейронов может вызвать и длительные ВПСП — до 80 мс. ВПСП сходной длительности можно зарегистрировать в таламических нейронах. Например, в нервных центрах мозга рыб, амфибий и рептилий ВПСП у различных нейронов также широко варьируют по длительности и форме и в ряде случаев они могут достигать большой длительности.
Механизмы генерации потенциала действия нервной клетки

Наличие трансмембранной разности потенциала нейрона может быть также объяснено неравномерным распределением неорганических ионов по обе стороны мембраны. При возбуждении возникает резкое повышение проницаемости мембраны к ионам натрия, поступление которых внутрь клетки и создает ПД. Остальные ионные механизмы деполяризации соматической мембраны, видимо, также сходны с таковыми для аксона. При возбуждении соответствующего афферентного нерва, пресинаптические окончания которого оканчиваются на данном нейроне (рис. 4, А, 2), или антидромном возбуждении аксона этого нейрона (рис. 4, А, 1), а также при деполяризации его сомы с помощью электрического тока (рис. 4, А, 3) нейрон генерирует ПД, амплитуда которого достигает 50—125 мВ. В большинстве нейронов ПД характеризуется теми же чертами, что и ПД нервного волокна, лишь длительность его для сомы нервной клетки несколько больше (от 1,5 до 3 мс). При всех этих способах активации нейрона на восходящей фазе (рис. 4, А) ПД сомы можно заметить «выемку». Для анализа этого явления было использовано антидромное раздражение мотонейрона при одновременной внутриклеточной регистрации его активности (рис. 4, А, нижний ряд). Было установлено, что в этом случае ПД состоит из трех отдельных компонентов, каждый из которых подчиняется закону «все или ничего». Экспериментально было доказано, что первый компонент (около 5 мВ) является ПД аксона (миэлинизированной части аксона). Второй (30—40 мВ) ПД немиэлинизированной части аксона, лежащего непосредственно у тела клетки, т. е. аксонного холмика. И третий компонент отражает активность сомы и дендритов нейрона. Для ПД аксонного холмика принят термин НС (потенциал начального сегмента аксона), для третьего СД (потенциал сомадендритной мембраны, рис. 4, Б). Причиной выемки, а иными словами, задержки перехода НС-потенциала в СД-потенциал является значительное увеличение площади поверхности мембраны в области аксонного холмика (переход к мембране сомы клетки). Это снижает гарантийный фактор дальнейшего распространения импульса. Пороговая поляризация для потенциала НС всегда ниже СД-потенципала и для мотонейрона составляет в среднем 10 мВ, в то время как для СД-потенциала — в среднем 30 мВ. Таким образом, при любой активации нейрона потенциал НС всегда предшествует СД-потенциналу. Для того, чтобы потенциал действия охватил сомадендритную мембрану, требуется некоторая ее критическая деполяризация, которая и создается потенциалом НС. Таким образом, различные участки мотонейрона по своим свойствам неоднородны. Сам импульс первично возникает не в теле клетки, а в аксонном холмике, переходя затем «антидромно» на сому. Такой механизм генерации ПД и общих чертах свойствен многим нервным клеткам, хотя есть и ряд особенностей.
Так, например, Кандел и Спенсер (1964), изучая механизм генерации импульсной активности в нейронах гиппокампа млекопитающих, нашли, что у 25% всех исследованных ими нейронов в основе возникновения ПД лежит более сложный процесс. Для этих нейронов характерно наличие еще одной зоны запуска ПД, предшествующей развитию ПД участка НС. В отличие от медленных ВПСП, предшествующих НС-потенциалу в мотонейроне, эта деполяризационная ступень была названа быстрым препотенциалом. (БПП) (см. рис.). Ими показано, что БПП представляет собой ПД, развивающийся по типу «все или ничего», а генерируется он в пределах дендритного дерева нейрона. Этот пиковый потенциал влияет на возникновение НС-потенциала и обычно ему предшествует. Поскольку у остальных 75% нейронов гиппокампа БПП отсутствует, то можно считать, что в гиппокампе преобладает обычная система возбуждения нейрона: НС—СД-потенциал. С наличием сходных локусов первичной зоны генерации ПД мы встретились, изучая внутриклеточную активность нейронов нервных центров амфибий и рептилий.
В нервных клетках различных животных могут быть сравнительно многообразные способы генерации ПД, однако наличие в нейроне звена НС—СД, видимо, носит более или менее универсальный характер.
Наличие низкопороговой зоны способствует интеграции всех деполяризационных и гинерполяризационных процессов, вызываемых в нейроне синаптической «бомбардировкой» как в покое, так и при действии афферентных сигналов, имеющих сигнальное значение. В связи с наличием такой зоны, которая находится на некотором отдалении от основной части сомы и дендритов нейрона, эффективность синапсов, вызывающих ВПСП на соме и дендритах, снижается в зависимости от степени их удаленности от НС. Не все ВПСП, особенно возникающие на удаленных от аксонного холмика участках мембраны дендритов, способны возбудить НС-потенциал. Иными словами, флюктуации ВПСП нейрона, регистрируемые внутриклеточно, могут не сопровождаться потенциалами действия клетки.
Итак, деполяризация, лежащая в основе ПД нейрона, имеет коренные отличия от деполяризации при ВПСП как по ионным механизмам, так и по важным физиологическим свойствам. Постсинаптическое возбуждение локально, градуально обладает способностью к суммации, в то время как нервный импульс является распространяющимся процессом и следует правилу «все или ничего». В связи с рефрактерностью процесс носит дискретный характер. Импульсы не сливаются друг с другом и не создают слитного деполяризационного процесса. Эти отличия в свойствах ВПСП и ПД были сгруппированы Грундфестом (1964). Он разделил мембрану нейрона на участки, электрически невозбудимые, но электрогенные (субсипаптическая мембрана), и электрически возбудимые, или мембраны с проводящим электрогенезом (внесинаптические участки нейрональной мембраны и особенно район начального сегмента аксона и вся аксональная мембрана).
Импульсная активность отличается от постсинаитического возбуждения (Вартанян, 1970) и по своему функциональному содержанию. ВПСП в нейроне действительно представляет собой процесс возбуждения в ответ на приходящую афферентную импульсацию. Импульсная же активность только передает на следующий нейрон или эффекторные органы в частотно закодированной форме суммарные амплитудные изменения мембранного потенциала нейрона, превышающие критический уровень деполяризации. Г. Я. Вартанян (1970) подчеркивает, что «если центральное возбуждение и торможение являются местными процессами, развивающимися в теле нейронов при активации соответствующих (возбуждающих или тормозящих) синаптических полей, то передача этих процессов (как возбуждения, так и торможения) и их взаимодействий осуществляется распространяющимися нервными импульсами, которые характером своего распределения во времени кодируют оба эти процесса в их взаимодействии и их количественные взаимоотношения в каждый данный отрезок времени».1
Механизмы фоновой или «спонтанной» активности нейронов
Хорошо известно, что большой процент нейронов ЦНС находится в состоянии постоянной импульсной активности. Для клеток различных отделов ЦНС характерен свой рисунок — паттерн спонтанной активности, который может быть более или менее постоянным или изменчивым. Внутриклеточные исследования показали, что такой активности нейронов обязательно предшествует деполяризация мембраны в виде быстрых препотенциалов или медленных волн разной формы.
Фоновая бомбардировка нейронов различных отделов мозга может либо непосредственно вызывать возбуждения нейрона, либо может создавать устойчивый уровень деполяризации, что вызывает ритмическую активность клетки (повторные ответы).
Механизмы фоновой активности нейрона могут объясняться, например, за счет синаптических воздействий, благодаря циркуляции импульсов в кольце соединенных друг с другом нейронов. О замкнутых цепочках нейронов говорил еще Лоренте де Но (1933). Кольцо может быть образовано и одним нейроном, за счет его возвратных коллатералей, оканчивающихся на соме или дендритах того же нейрона. Такое возвратное возбуждение может, видимо, осуществляться и через специальные вставочные нейроны. Так, например, Асанума и Брукс (Asanuma, Brooks, 1963) на пирамидных нейронах кортикоспинального тракта нашли, что антидромное раздражение послед них может вызвать возвратное возбуждение последних в виде короткого (около 40 мс) повышения возбудимости.

Б — возвратного торможения; В, Г — различных способов пресинаптического торможения; Д — прямого торможения, Е — торможения (без вставочного нейрона в зрительной коре) (по Ватанабе и др.).
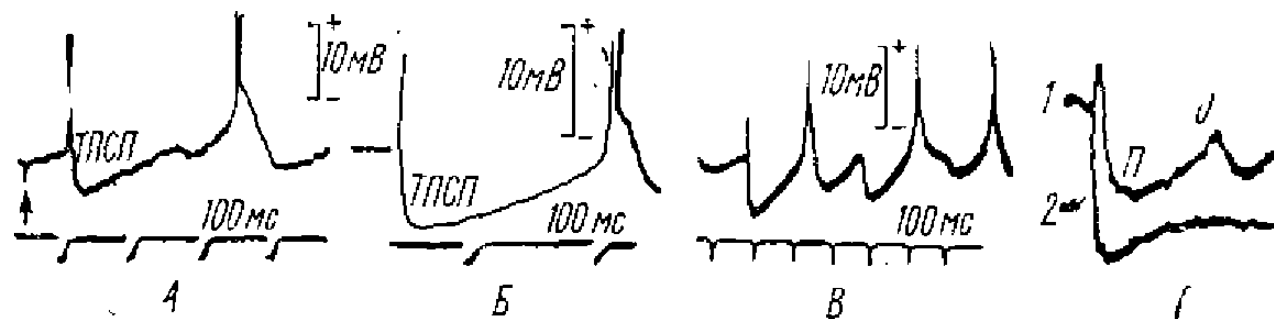
Эти результаты были уточнены в опытах с внутриклеточной регистрацией потенциалов нейронов пирамидного тракта при их антидромной активации (Stefanis, Jasper, 1964). Было выяснено, что после прихода антидромного импульса в нейрон вначале наблюдается длительный, более 100 мс, тормозный постсинаптический потенциал (ТПСП). Часто перед этим тормозным потенциалом наблюдался деполяризационный потенциал меньшей длительности и с более коротким латентным периодом, который имел все характеристики ВПСП. Видимо, он вызывался через возвратные коллатерали и вставочные нейроны. Наличие возвратных возбуждающих связей в зрительной коре больших полушарий кролика и обонятельной луковице лягушки было показано также и в нашей лаборатории (Гусельников, Супин, 1968; Гусельникова, Гусельников, 1975), Схема возвратного возбуждения показана на рис. 5, А.
Периодичность (ритмичность) спонтанной активности нейронов может определяться и синхронным развитием в нейронах ТПСП в связи с активностью вставочных тормозных нейронов. И, наконец, возникновение фоновой активности может быть связано со спонтанным выделением медиатора, и особенно в тех случаях, когда синапсы расположены на тонких разветвлениях дендритов, обладающих высоким RBX.
В интегративных функциях, свойственных ЦНС, нейрон представляет наименьшую интегрирующую единицу, которая способна «оценивать», «сравнивать», суммировать и преобразовывать приходящие сигналы. Более крупными интегрирующими единицами являются нейронные ансамбли (сети). Так, Берль (Beurle, 1956) считает, что такие ансамбли формируются из близко расположенных нейронов, которые способны к синхронизации своей активности благодаря ряду специальных механизмов. А. Б. Коганом и его сотрудниками показано, что такая синхронизация активности нейронов возможна в пределах 300 мкм. В синхронизации же соседних элементов кроме синаптических механизмов играют, видимо, роль и электротонические влияния, а также внеклеточные электрические ноля, создаваемые самими нейронами (Bennet et al., 1963; Эзрохи, 1967; Русинов, 1969; Eckert, 1963, и др.). В основе синхронизации могут лежать также изменения обмена веществ в ансамблях, нейронов, гормоны, продукты метаболизма и т. д. Известно, что наибольшая синхронизация наблюдается при одновременной синаптической активации нейронной системы. Уже исходя из этого, как мы увидим далее, необходимо, видимо, признать, что синхронная активация нейронных ансамблей не должна зависеть от расстояния, если она связана со специальными пейсмекерами — водителями ритма. В этом случае синхронизация должна охватывать огромное число нейронных ансамблей на разных уровнях головного мозга. По всей видимости, наличие в ЦНС различных способов и путей синхронизации нейрональной активности является важным и необходимым фактором в интегративной нервной деятельности.
Торможение нервных клеток
На нейронном уровне обнаружено два основных вида торможения в центральной нервной системе — пресинаптическое и постсинаптическое.
Пресинатическое торможение (ПТ) осуществляется посредством специфической синаптической деполяризации возбуждающих пресинаптических окончаний посредством аксо-аксональных синапсов. Это уменьшает распространяющийся по окончаниям ПД и резко уменьшает (или блокирует) выход возбуждающего медиатора из пресинаптического окончания в синаптическую щель, что и проявляется в подавлении ВПСП.
Пресинаптическую деполяризацию связывают сейчас со сложным механизмом. Предполагается, что путь ПТ является полисинаптическим и что аксо-аксональные синапсы образуются особой группой вставочных нейронов, оказывающих деполяризующее действие на пресинаптические окончания (см. рис. 5, В, Г).
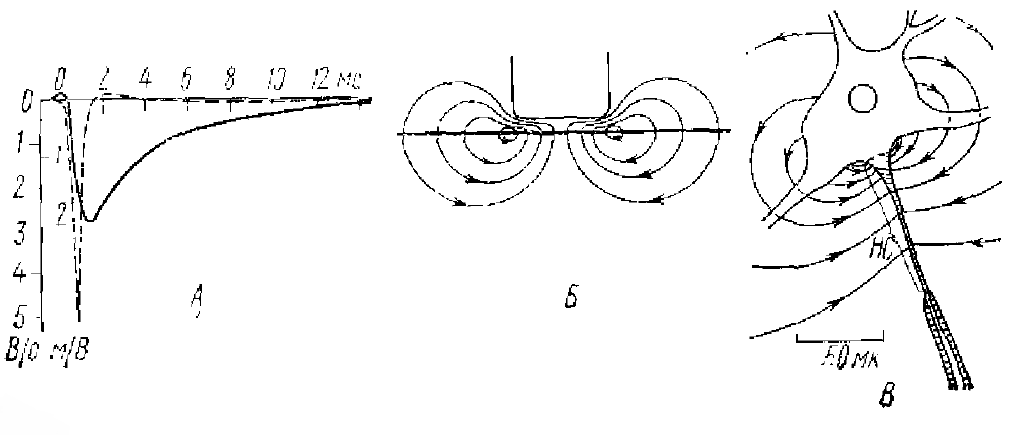
Второй способ торможения нервных клеток — постсинаптическое торможение, которому соответствует процесс гиперполяризации в постсинаптической мембране нейрона. Он был назван тормозным постсинаптическим потенциалом (ТПСП). Развитие ТПСП также связано с изменениями ионной проницаемости субсинаптических участков мембраны нейрона. По своим ионным механизмам оно значительно отличается от генерации ПД и ВПСП. Не разбирая подробно ионные механизмы ТПСП, можно кратко отметить, что «если возбудительные синапсы стремятся свести мембранный потенциал к нулю, открывая натриевые и калиевые «поры» в субсинаптической мембране действием возбуждающего медиатора, то тормозные синапсы, наоборот, стремятся довести мембранный потенциал приблизительно до 80 мВ (для мотонейрона), открывая калиевые и хлорные «поры» тормозной субсинаптической мембраны. Если в случае возбудительного синапса (см. рис. 2, Б) ток от остальной части постсинаптической мембраны направлен по калиево-натриевым «порам» внутрь, то в случае тормозного синапса — по хлорно-калиевым — наружу» (рис. 6, Б) (Экклс, 1971). Таким образом, торможение активности мембраны клетки есть, по сути дела, процесс, направленный на затруднение развития процессов деполяризации.
Для ТПСП характерна большая длительность, чем для ВПСП. В основе значительной длительности ТПСП у многих нейронов и особенно у многих нейронов высших этажей ЦНС лежат разные причины: временная дисперсная активация различных тормозных синапсов, свойства субсинаптической мембраны, свойства вставочных нейронов и ряд других причин. Так, например, длительные ТПСП возникают в результате ритмического разряда, тормозных клеток Реншоу, который вызывается импульсами, идущими по коллатералям моторного аксона. Наиболее мощные ТПСП имеют большую длительность, что связано с частотой ритмического разряда тормозного нейрона, которая доходит до 1000 имп/с. ТПСП нейронов головного мозга позвоночных были подвергнуты наиболее интенсивному изучению в последние годы. Полученные данные свидетельствуют о том, что почти все ТПСП этих нейронов (рис. 7) имеют длительность 100 мс и более.
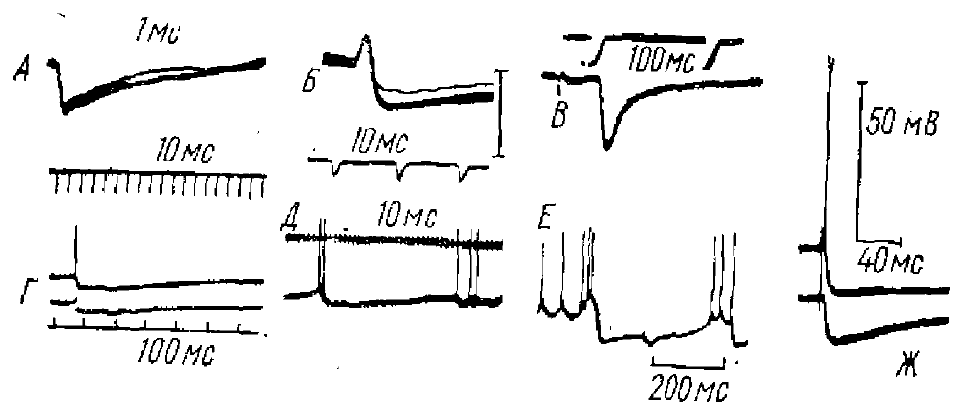
ТПСП длительностью 20 мс более редки. В таламических нейронах млекопитающих, нейронах клиновидного ядра и ряде других нейронов ТПСП достигает 100 мс и более. Большой амплитуды и длительности достигают ТПСП в пирамидных клетках гиппокампа млекопитающих — порядка 200 мс. Такой же длительности достигают ТПСП и в пирамидных корковых нейронах (рис. 7, Г). В исследованиях показано широкое разнообразие ТПСП в нейронах различных нервных центров рыб, амфибий и рептилий, где они могут достигать как очень большой длительности, так и быть более короткими, порядка 15—20 мс (например, у нейронов обонятельной луковицы рыб, амфибий рептилий).
Особенности строения тормозных синапсов
Поскольку функционально для ряда синапсов удалось точно установить их тормозную природу, стало возможным использование электронной микроскопии для изучения особенностей их строения. Греем (Gray, 1959) и Хемлином (Hamlyn, 1962) была сделана попытка выделить отличительные признаки в строении тормозных синапсов по сравнению с возбудительными. Полагают, что для тормозных синапсов характерна более узкая синаптическая щель (200 ммк) и менее плотная и толстая субсинаптическая мембрана. Если у возбуждающих синапсов зона уплотнения занимает большую часть противолежащих синаптических мембран, то у тормозных эта зона ограничена небольшими участками. Однако открытие синапсов промежуточной структуры подвергает сомнению такую классификацию. Возможно, более удачным является идентификация тормозных синапсов по величине и форме везикул (синаптических пузырьков). Считается, что в отличие от возбудительных синапсов, имеющих круглые везикулы, везикулы в тормозных синапсах имеют овальную форму, они более мелкие и имеют уплощенный вид. Насколько эти критерии точны и универсальны для тормозных синапсов, покажут лишь будущие исследования.
Большинство исследователей признают существование двух различных типов синапсов: возбуждающих и тормозящих, что связывается обычно с химическими свойствами пресинаптических аксональных окончаний. В связи с этим предполагается наличие специализированных тормозящих нейронов, пресинаптические окончания которых выделяет особый медиатор, оказывающий тормозящее действие на последующий нейрон. Ряд исследователей считает, что дело в особых свойствах субсинаптических мембран, т. е. именно с этими мембранами связаны различные эффекты действия одного и того же медиатора.
Экклс (1971) пытался проанализировать этот вопрос и склонялся к двум общим принципам, сформулированным Дейлом (1935) для синапсов с химической передачей. Первый принцип Дейла заключается в том, что все терминали одного нейрона выделяют один и тот же медиатор. Второй — говорит о том, что медиатор во всех терминалях одного нейрона открывает ионные поры одного типа, свойственного тормозным или возбудительным синапсам.
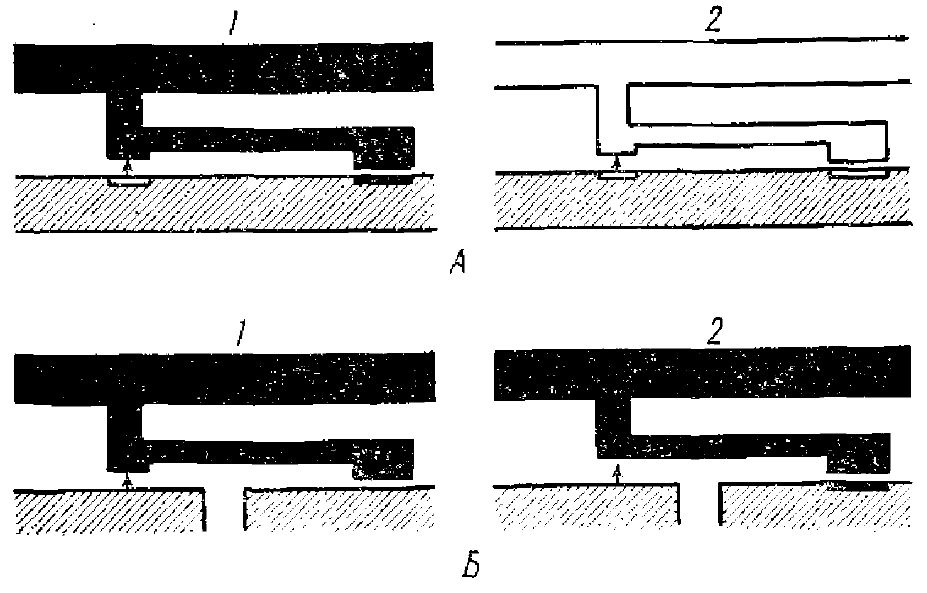
Экклс считает, что нейрон на стадиях формирования своих отростков обладает или тормозной или возбудительной специфичностью. Он полагает (рис. 8, А, 1, 2), что при развитии коллатералей тормозного нейрона они, в силу своей химической чувствительности, находят сами участки на поверхности нейрона, которые специализировались как тормозные (рис. 8, Л), отвергая возбудительные синаптические участки. В этом случае образуется тормозной синапс (рис. 8, А, 1). То же наблюдается и со специализацией возбудительных синапсов (рис. 8, А, 2).
Второй вариант, по Экклсу, состоит в том (рис. 8, Б, 1, 2), что уже специализированные коллатерали аксона, например тормозные, обладают чувствительностью к определенным участкам поверхности нейрона, но еще «химически» не специализированным (рис. 8, Б, 1) и только потом сами создают на субсинаптической мембране специфическое для тормозного синапса свойство мембраны (рис. 8, Б, 2).
Особый интерес представляют дендро-дендритические синапсы, обнаруженные уже сейчас во многих мозговых образованиях млекопитающих. Интересно отметить, что учитывая только электронно-микроскопические данные, о характере уплотнения мембран и расположения синаптических пузырьков, такой синапс является возбуждающим по отношению дендрита одного нейрона и тормозящим по отношению дендрита другого нейрона. Какова медиаторная природа таких синапсов, пока не известно.
В последние годы электронная микроскопия значительно расширила наши знания о диапазоне способов контакта одних нервных клеток с другими. Так, например, показано наличие сома-дендритических и дендро-соматических синапсов в обонятельной луковице, в верхних слоях tectum opticum среднего мозга лягушки и ряд других интересных синаптических контактов. Возможно, что своеобразие такой структуры синапсов будет хорошо коррелировать с основными принципами Дейла, но не исключены и отклонении от этих правил.
Распределение тормозных и возбуждающих синапсов на теле и отростках клетки и их функциональные особенности. Для ряда нейронов ЦНС экспериментально показано, что тормозные синапсы обычно расположены на телах нейронов (а у некоторых даже на аксонном холмике): пирамидных клеток гиппокампа, клеток Пуркинье мозжечка, клеток-зерен зубчатой фасции гиппокампа, пирамидных клеток коры больших полушарий, мотонейронов, клеток таламо-кортикального реле. Однако тормозные синапсы, расположенные на дендритах нейронов, были найдены для клеток-зерен и клеток Пуркинье мозжечка, для митральных клеток обонятельной луковицы. Что касается возбуждающих синапсов, то они могут располагаться как на теле клетки и аксонном холмике, так и на базальных и апикальных дендритах. Однако у разных нейронов головного мозга позвоночных место и плотность их расположения широко варьирует. Пространственное расположение возбуждающих и тормозящих синапсов на нейроне является основным фактором взаимодействия всех де- и гиперполяризационных влияний и играет основную роль в интегративной деятельности.
Суммарная биоэлектрическая активность мозга должна отражать какие-то сложные электрические процессы одновременно, более или менее синхронно возникающие в относительно большой популяции нейронов. Поэтому, как бы ни были важны для нас знания о пространственно-временном течении процессов возбуждения и торможения на уровне одного нейрона, нас прежде всего должны интересовать те механизмы нейрональной деятельности, которые обуславливаются связями в нервной сети между различными нейронами, играющими главную роль в их одновременной, синхронной работе.
Морфологические исследования, которые привели к открытию сложнейших нейрональных систем мозга, впервые поставили вопрос о механизмах их работы. И на первых этапах как физиологи, так и морфологи делают попытки построения таких функциональных систем, опираясь на возможные гипотетические и большей частью умозрительные схемы о назначении той или иной сложноорганизованной сети нейронов. Исходя из таких систем, было выдвинуто значительное количество теорий и гипотез о механизмах работы мозга. Естественно, что с применением микроэлектродной техники и вскрытием основных закономерностей работы нервных клеток и механизмов их связей эти теории сейчас носят в своем большинстве исторический интерес. Все эти схемы опирались на представление только о возбудительной природе импульсации, и, как отмечал Экклс (1971), им был свойствен основной недостаток — отсутствие знаний о наличии синаптического тормозного действия.
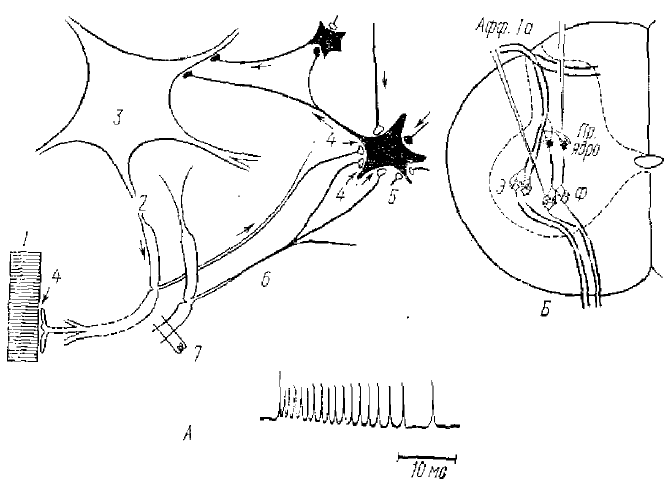
Впервые в работах Реншоу (Ranshaw, 1941) было показано наличие в ЦНС специфических тормозных синапсов. Разряд импульсов в моторных аксонах спинного мозга затормаживает все типы мотонейронов, которые расположены на данном сегментарном уровне. Такое торможение легко вызвать электрическим антидромным раздражением мотонейрона. Оно связано с тем, что аксоны мотонейронов дают несколько коллатералей, которые образуют синапсы на вставочных нейронах и, активируя их, вызывают длительные, высокочастотные разряды. Эти вставочные нейроны, получившие имя исследователя,— клетки Реншоу, оказывают тормозящие действия на мотонейрон (рис. 9, А), вызывая в нем длительные ТПСП. Естественно, что такое торможение в норме возникает в мотонейроне и рефлекторно. Наиболее изучен так называемый прямой тормозной путь к мотонейронам по волокнам группы 1 а (рис. 9, Б). В этом случае импульсы, идущие от аннуло-спиральных окончаний в мышечных веретенах той или иной мышцы, оказывают тормозное действие на мотонейроны мышцы антагониста через клетки Реншоу. Такое торможение было названо прямым торможением. Его называют и первичным торможением, т. к. оно связано с активацией особых тормозящих структур и для клетки является первичным процессом (т. е. оно не связано с предварительным ее возбуждением). Экклс (1971) называет это торможение поступательным (см. рис. 5, Б).
Говоря об антидромной активации, указывалось, что аксоны мотонейронов дают ряд коллатералей, оканчивающихся на клетках Репшоу; в свою очередь коллатерали клетки Реншоу могут оканчиваться тормозными синапсами на многих мотонейронах. Таким образом, в случае возбуждения мотонейрона спайковый потенциал, распространяясь по его аксону, заходит в коллатерали, возбуждая вставочный тормозной нейрон. В результате мотонейрон, возбужденный мгновенье назад, затормаживается,— происходит самоторможение нейрона. Это торможение получило название возвратного торможения, при коллатерального постсинаптического торможения (см. рис. 9, А). Схема такого торможения приведена на рис. 5, Б. И, наконец, третьим возможным способом торможения является пресинаптическое торможение, о котором мы говорили ранее (см. стр. 35) (рис. 5, В, Г).
Ватанабе и др. (Watanabe et al., 1966) предполагали наличие еще одного способа торможения головного мозга для корковой зоны зрительного анализатора за счет внутрикортикальных моносинаптических (т. е. без вставочного нейрона) коллатеральных тормозных путей (см. рис. 5, Е). Иными словами, выдвигалось предположение, что все нейроны, которые первично возбуждаются, имеют аксонные коллатерали тормозного действия, вызывая торможение но механизму обратной отрицательной связи, а по своим главным аксонным ветвям эти же нейроны оказывают возбудительное действие. Однако эта схема крайне гипотетична и не имеет пока экспериментальных доказательств.
Вторичное торможение нейрона есть, по сути дела, следствие его возбуждающей активации, т. е. возбуждение нейрона само по себе подавляет последующее его возбуждение. В ряде нейронов, например, ретикулярной формации ствола, коры больших полушарий, промежуточных нейронах спинного мозга синаптическая деполяризация оказывается очень стойкой и потенциал действия не ослабляет и не устраняет остаточное синаптическое действие, а сам потенциал действия не сопровождается последующей выраженной гиперполяризацией. В результате этого на фоне ВПСП возникает, как правило, ритмический спайковый разряд высокой частоты. При сильной синаптической активации в таких нейронах может наступить частичное или полное торможение спайковой активности в связи с чрезмерной синаптической деполяризацией мембраны. Такая высокая деполяризация мембраны может быть связана с процессами, затрудняющими инактивацию или выведение медиатора из синаптической щели, что поддерживает мембрану длительное время в активированном состоянии. Особенно это может наблюдаться при ритмической полисинаптической активации, когда развивается стойкая деполяризация постсинаптической мембраны, превосходящая критический уровень для генерации потенциала действия. Вторичное торможение может возникнуть и за счет блокады пресинаптических окончаний (отличного от пресинаптического торможения) при высоких частотах пресинаптического возбуждения. В этом случае в пресинаптических окончаниях блокируется проведение нервных импульсов и прекращается их транссинаптическое действие.
Пути торможения в некоторых структурах головного мозга позвоночных
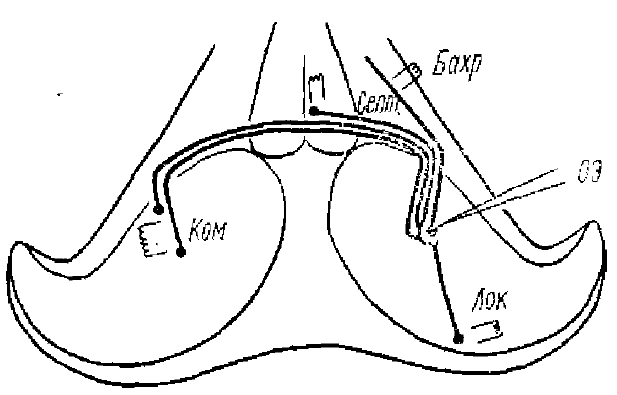
Андерсену и др.. 1964)
В качестве примера коркового типа возвратного торможения можно привести данные Андерсена и др. (Andersen et al., 1963, 1964), полученные на нейронах гиппокампа. Было показано, что раздражение различных афферентных путей гиппокампа вызывает в пирамидальных клетках большие (до 20 мВ) и длительные (до 200 мс и более) ТПСП (рис. 10). Волокна всех раздражаемых путей конвергируют на пирамидных клетках, от которых ведется регистрация.
Поскольку все виды раздражения вызывают тормозные постсинаптические потенциалы почти во всех пирамидных клетках гиппокампа, можно было предположить наличие опосредующего, вставочного нейрона для такого эффекта торможения. Этот нейрон должен обладать следующими свойствами: 1) его аксон широко ветвиться и оканчиваться на многих пирамидных клетках, 2) синаптические окончания должны быть расположены на соме пирамидных клеток и 3) он должен активироваться при любом из указанных выше способов раздражения, или косвенно, т. е. через коллатерали аксонов пирамидных клеток после их возбуждения.
Гипотетические тормозные нейроны авторы пытались определить как анатомически, так и физиологически. Так, вставочным нейроном с анатомической точки зрения могут быть корзинчатые клетки, поскольку аксон каждой из них широко ветвится и образует синаптические контакты с сомой 200—500 пирамидных клеток (рис. 11). На рис. 11 видно, что корзинчатая клетка получает возбуждающие синапсы от коллатералей аксонов пирамидных клеток, а сама образует много тормозных синаптических окончаний на телах пирамидных клеток. Необходимо подчеркнуть, что локализованные на соме пирамидных клеток тормозные синапсы занимают самое оптимальное положение для регуляции возникновения импульсных разрядов сомы пирамидных клеток, т. к. они расположены между возбуждающими синапсами на дендритах и аксонным холмиком, где генерируется импульс.
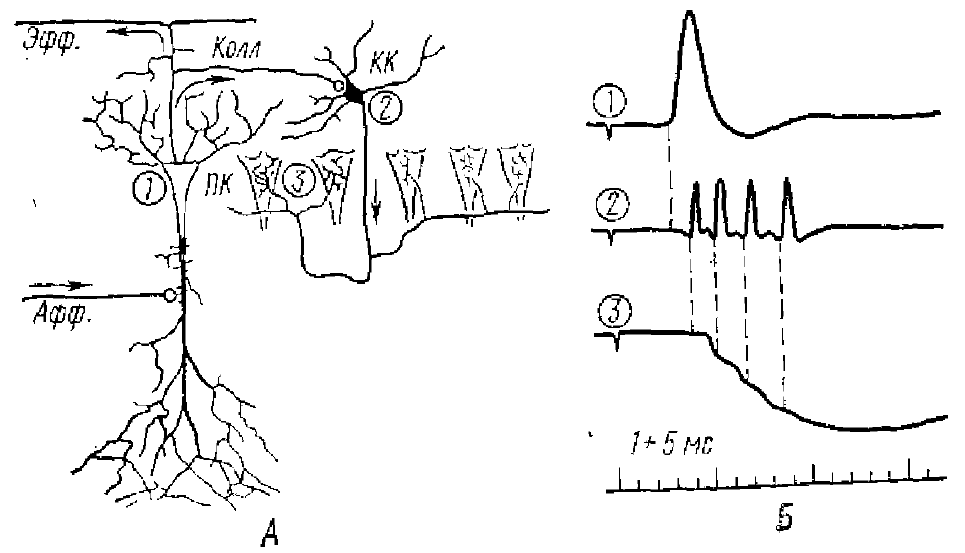
В физиологических опытах тормозные нейроны также были найдены. На уровне залегания именно корзинчатых клеток была записана активность, характерная для тормозного нейрона и заключающаяся в высокочастотном его разряде до 500-1000 имп/с, т. е. приблизительно таким же, как и в клетках Реншоу, Особенность этой клетки гиппокампа, в отличие от его пирамидной клетки, состоит в том, что она активируется при любом из применяемых видов стимуляции. Клетка обнаруживает длительный ВПСП с наложенными на пего спайками, без каких-либо признаков ТПСП.
Отличительной особенностью гиппокампа является значительное преобладание тормозных процессов, которые могут быть зарегистрированы даже при очень слабых раздражениях. Действительно, ТПСП в пирамидном нейроне можно зарегистрировать даже без его разряда (рис. 12, Б, 3). ТПСП могут возникать в результате непосредственного возбуждения корзинчатых клеток афферентным залпом (Андерсен, Ломо, 1967). Такое торможение было названо опережающим торможением.
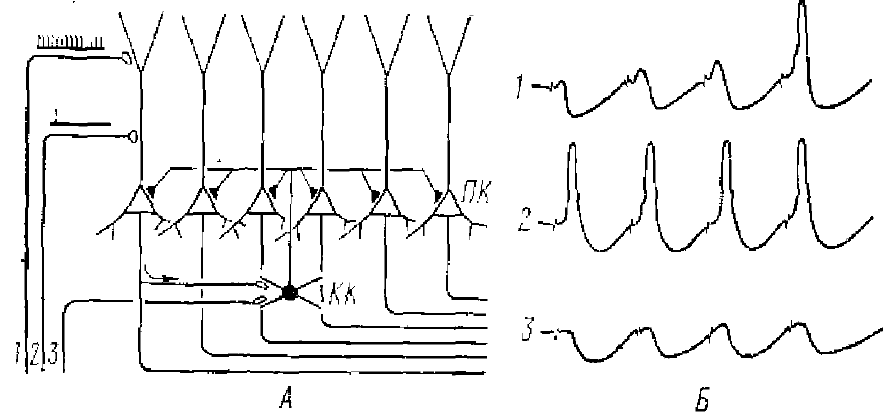
Возвратное торможение (Andersen et al., 1966) характерно и для другой зоны гиппокампа — area dentata.
Механизм возвратного торможения для таламических образований млекопитающих
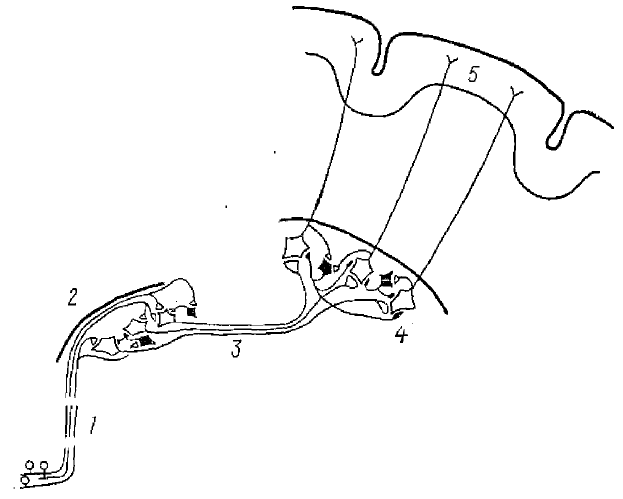
В исследованиях Андерсена, Брукса, Сиерса и др. возвратное торможение было показано на примере работы нервной сети вентробазального ядра таламуса, являющегося таламо-кортикальным реле (ТКР) передача сенсорной информации. На рис. 13 показан путь к сенсорной коре для волокон кожного нерва передней конечности. Торможение в клиновидном ядре является прямым (или поступательным), а в вентробазальном ядре — возвратным. Правильность этой схемы была подтверждена экспериментально. Большие и длительные ТПСП вызываются в нейронах вентробазального ядра таламуса либо ортодромными импульсами, возникающими при раздражении локтевого и медиального нервов, либо антидромными импульсами, возникающими при раздражении корковых окончаний таламо-кортикальных волокон (рис. 14). Причем ТПСП могут возникать независимо от того, дает ли данный нейрон ПД или нет. ТПСП наблюдается во всех нейронах вентробазального ядра, аксоны которых идут к коре головного мозга. Таким образом, через возвратный тормозной путь (Т-клетка) разряд таламических нейронов вызывает мощное и широко распространяющееся торможение, которое подавляет все таламические разряды на 100 мс и более, причем ТПСП возникают и в тех нейронах, которые не давали перед этим ПД, так как, видимо, имеет место широкий охват тормозными коллатералями вставочного нейрона многих нейронов ТКР (рис. 15). При регистрации активности от нейронов вентробазального ядра таламуса Андерсеном, Экклсом и Сиерсом (1964) были найдены такие предполагаемые тормозные клетки, хотя морфологически они пока и не идентифицированы.
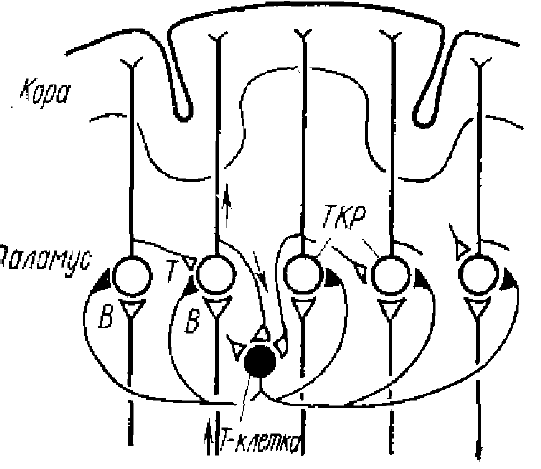
Экклс (1971) полагает, исходя из данных Рамон-и-Кахала, что тормозными вставочными нейронами в ТКР могут быть клетки типа Гольджи II — крупные клетки с ветвящимися аксонами и многочисленными ветвями дендритов. Эти нейроны, видимо, могут иннервироваться коллатералями, отходящими от аксонов клеток ТКР, а множественные короткие ветви аксонов клеток Гольджи II образуют синапсы на многих клетках ТКР.
Система возвратного торможения найдена и для другого таламического образования — наружного коленчатого тела (НКТ) (Burke, Sefton, 1966, и др.). При внутриклеточной регистрации от нейронов НКТ было показано, что как при одиночном раздражении зрительного нерва, так и при раздражении зрительной коры возникают ТПСП так же, как и в нейронах вентробазального ядра таламуса. При стимуляции зрительного нерва было выяснено, что в нейронах коленчатого тела латентный период ВПСП был короче, чем ТПСП. На основании того, что разность составляет 1 мс, был сделан вывод, что она определяется наличием в цепи проведения вставочного тормозного нейрона.
Принцип возвратного торможения свойствен, видимо, всем трем функциональным группам ядер таламуса млекопитающих — специфическим, неспецифическим и ассоциативным. Возвратное торможение характерно и для различных нервных центров домлекопитающих позвоночных. Так. в обонятельной луковице система возвратного торможения обнаружена у всех исследованных нами классов позвоночных: рыб, амфибий, рептилий (Гусельникова, Гусельников, 1975). В последнее время нами найдено возвратное торможение и для нейронов первичной коры амфибий, и для нейронов коры переднего мозга рептилий. Во всех этих случаях возвратное торможение опосредуется интернейроном.
Что касается коры больших полушарий, то Экклс дает две возможные гипотетические схемы возбудительных и тормозных связей — одну для синаптических связей аксонных коллатералей пирамидных клеток (рис. 1В), другую для связей специфических афферентных волокон с пирамидными клетками коры (рис. 17).
На основании того, что антидромная активация пирамидного тракта приводит в пирамидных нейронах к ТПСП, было сделано заключение о наличии в коре вставочных тормозных клеток. В одних клетках латентный период ТПСП точно соответствовал задержке «равной» одному вставочному нейрону, в других — между приходом антидромного импульса в кору и началом ТПСП происходила большая задержка. Было сделано предположение, что такая задержка объясняется более длинными полисинаптическими путями, которые включают вставочные нейроны. На рис. 16 и 17 видно, что такими вставочными нейронами являются звездчатые клетки, причем они могут быть как возбудительными, так и тормозными. Исходя из гистологических данных, предполагают, что некоторые из них могут быть тормозными клетками, которые образуют у пирамидных клеток корзинчатые образования типа аналогичных образований у пирамидных клеток гиппокампа. Однако до сих пор еще нет точных доказательств этого положения. Предполагается также, что коллатерали аксонов пирамидных клеток имеют небольшое количество прямых связей с тормозными нейронами, а большей частью торможение осуществляется посредством полисинаптических путей. Известно, что ТПСП в пирамидных нейронах коры возникают и при афферентных воздействиях при раздражении специфических и неспецифических путей, ведущих к коре.
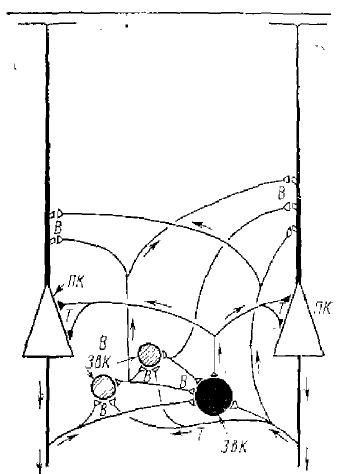
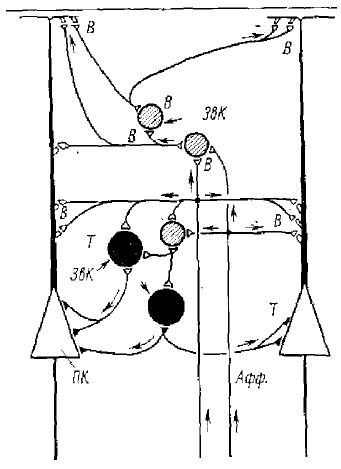
Однако до сих пор еще не ясно, осуществляется ли это тормозное действие за счет разрядов самих пирамидных нейронов с последующей активацией аксонных коллатералей (см. рис. 16) или за счет тормозных путей, активируемых афферентными путями (см. рис. 17). Принципиально обе эти схемы не различаются, хотя пока крайне гипотетичны, поскольку вопрос об идентификации тормозных нейронов коры еще не решен.2 Сложная картина путей возбуждения и торможения в головном мозге описана и для нейронов мозжечка, где также найдено и прямое и возвратное торможение (Экклс, 1971),
Таким образом, можно считать, что нервные центры самых различных уровней организации и во всем ряду позвоночных животных имеют некоторые общие принципы организации процессов возбуждения и торможения в мозге, хотя конкретные пути и интимные механизмы этих процессов и их пространственно-временные характеристики определяются спецификой структуры и функциональными особенностями нейронов и их связей для каждого конкретного нервного центра.