В последние 15—20 лет многочисленными исследованиями ряда авторов было показано, что суммарная электрическая активность коры мозга не исчерпывается диапазоном электрических колебаний, формирующих ЭЭГ, а включает потенциалы большей длительности, период колебания которых может достигать минут и часов.
Длительные электрические потенциалы в виде медленных сдвигов уровня постоянного потенциала и его колебаний, регистрируемые в коре мозга, были предметом изучения как отечественных (Аладжалова, 1960, 1962; Русинов, 1955; Шуранова, 1966; Швец, 1958, 1963), так и зарубежных исследователей (Morell, 1961; Goldring, O’Leary, 1951, 1954; Arduini et al., 1957; Bures, 1957, и многие другие).
Постоянный потенциал коры головного мозга (ПП) и его механизмы
Этот потенциал известен так же как квазипостоянный потенциал, потенциал постоянного тока — «ДС-потенциал», стабильный потенциал, постоянная составляющая ЭЭГ.
Фоновый градиент ПП—хорошо установленный факт для многих нервных структур, в том числе и для коры больших полушарий; что же касается его генеза, то этот вопрос более сложен. ПП регистрируются с разных участков мозга. Своим происхождением они обязаны ионным перепадам, возникающим в результате различия обменных процессов в разных структурах, разной полярности различных частей нейрона, разности ионных концентраций на тех или иных анатомических границах. Так как такая разность потенциалов без каких-либо специальных воздействий сохраняется длительное время на постоянном уровне, ее и назвали постоянным потенциалом.
Такие градиенты постоянного потенциала в коре больших полушарий могут носить поперечный, продольный и трансколлазальный характер.
Наиболее распространенной точкой зрения на источник стабильной разности потенциалов является гипотеза Либет и Жерар (Libet, Gerard, 1941), поддержанная Голдрингом и О’Лири (Goldring, O’Leary, 1951), в которой постулируется, что ПП создается благодаря поляризации и одинаковой (радиальной) направленности нейронов.
Другая гипотеза о происхождении ПП была высказана Чирджи и Тэйлором (Tschirgi, Taylor, 1958). Они полагают, что стойкая разность потенциалов наступает вследствие распределения ионов на гематоэнцефалическом барьере, так как между электродом, введенным в в вену, и поверхностью коры мозга эта разность составляет 1—5 мВ; причем кора по отношению к венозной крови электроотрицательна. Далее было показано, что эта разность потенциалов существует между различными отделами мозга и рядом неактивных структур (костной тканью, мозговой жидкостью и т. д.).
Кроме этих гипотез существуют и другие точки зрения на генез ПП. Ряд исследователей полагает, что ПП могут являться результатом суммации ПСП нейронов (так называемых медленных ПСП). Экклс и сотрудники (Eccls et а]., 1962) связывали генез ПП с длительными электротоническими потенциалами в связи с пресинаптической деполяризацией, Хилд и др. (Hild et al., 1958) — с мембранным потенциалом глиальных клеток и т. д. Однако до сих пор этот вопрос окончательно не решен, хотя участие как нейронов, так и глии в его генезе несомненно.
ПП между разными точками коры по отношению желудочка обнаружены как у животных, так и у человека. У человека ПП были обнаружены и в глубоких структурах мозга. Кроме того, существует определенный градиент ПП между различными точками на поверхности коры, а также между разными слоями коры.
Более правильной и прямой проверкой гипотезы Либета и Жерара явилось бы измерение мембранного потенциала одного пирамидного нейрона коры в разных его участках, что для новой коры пока методически невозможно. Поэтому используется обычная проходка коры вертикально ориентированным экстраклеточным электродом.
Н. А. Аладжалова и О. X. Коштоянц (1957) при помощи неполяризующихся внеклеточных микроэлектродов показали, что при погружении электрода в глубь коры мозга, потенциал сдвигался в сторону негативности и на уровне V—VI слоя достигал максимума. Они сделали вывод, что в верхних и нижних слоях коры мозга возникает электрическое поле противоположного знака (рис. 1, 2). Экстраклеточные токи в верхних слоях коры мозга в этом случае должны быть направлены к соме нейрона. Авторы связывают этот потенциал с нейронами, воспринимающими передачу возбуждения по неспецифическим путям. Возникновение таких экстраклеточных токов может, видимо, приводить к изменению возбудимости нейрона путем де- или гиперполяризующего их действия на мембрану.
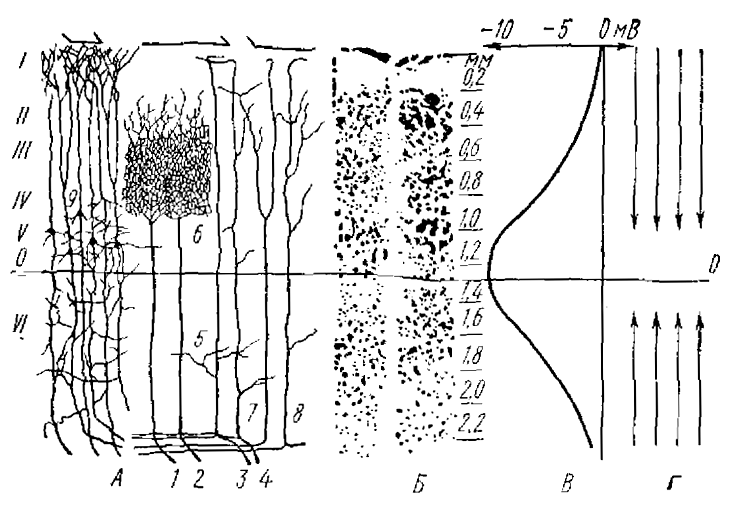
I—VI — слои коры; 1—2 — специфические афферентные волокна из таламуса. 3—4 — неспецифические афферентные волокна, 5 — аксо-дендритические, 6 — аксо-соматические окончания афферентных волокон, 7—8 — ассоциативные волокна, 9 — крупные пирамидные клетки с длинным аксоном

По оси ординат — разность потенциалов между поверхностью коры полушарий и глубоким слоем коры мозга, по оси абсцисс — глубина погружения микроэлектрода. Толщина коры мозга 2 мм, далее микроэлектрод погружался в белое вещество
Другие исследователи (Tschirgi, Taylor, 1958) не нашли никаких изменений ПП при погружении электрода в кору, если записывалась разность потенциалов между последней и кровяным руслом, на основании чего они отрицают роль корковых нейронов в генерации ПП коры. Однако эти данные скорее показывают, что дипольные структуры коры не единственный источник ПП (Шуранова, 1966).
ПП коры претерпевают изменения — апериодические сдвиги, при действии самых различных факторов. Величина и полярность ПП прямо зависит от функционального состояния животного и от условий опыта; наркоз, изменение температуры тела и pH крови, сон могут значительно изменять исходные градиенты ПП. Сдвиги ПП более чувствительны к условиям опыта, чем ЭЭГ или вызванные потенциалы (Русинов, 1969; Brookhart et al., 1958; Gumnit, 1961, и др.). Значительное влияние на ПП оказывает электрическое раздражение коры — ее поляризация. Так, при поляризации поверхности коры катодом уровень ПП сдвигается в сторону негативности, при поляризации анодом — в сторону позитивности (Goldring, O’Leary, 1951; Gaspers, 1962, и др.).
Однако в лаборатории В. С. Русинова, Г. Д. Кузнецовой (1963) было показано, что величина отклонения ПП и его знак на действие поляризующего тока зависят от исходного уровня ПП коры. Чем более позитивен исходный ПП, тем он больше негативируется при действии анода; при негативном уровне ПП происходят позитивация на ток. Если же ПП равен 1—2 мВ, то он наиболее стабилен и меньше сдвигается внешними воздействиями.
При тетаническом раздражении поверхности мозга импульсами тока значительной интенсивности, с частотой 50-100 в 1 с, возникает длительный отрицательный потенциал, который связывают с дендритами, хотя его генез ряд исследователей трактуют по-разному (Purpura,1933; Ройтбак, 1969, и др.). На одиночный удар тока, приложенный к поверхности коры, возникает, как мы уже говорили, дендритный потенциал, а затем медленный отрицательный потенциал (МОП), генеза которого мы коснемся несколько позже.
➥ Более подробно: Физиология дендритов нервных клеток и их функциональная роль
Сдвиги ПП коры наблюдаются и при раздражении различных подкорковых структур мозга. При высокочастотном раздражении ретикулярной формации (РФ) ствола вместе с эраузал-реакцией наблюдается сдвиг ПП в сторону негативности с крутым фронтом нарастания и медленным его спадом в течение ряда секунд (Arduini et al., 1957; Kanai, Katzman, I960; Vanasupa et al., 1959).
Что касается генеза этих сдвигов ПП при раздражении РФ ствола (так же как и при раздражении РФ среднего мозга, Arduini et al., 1957), то мнения исследователей расходятся: одни относят их за счет нейрональных структур коры, другие — за счет ненейрональных структур (глии или гематоэнцефалического барьера).
Значительный экспериментальный материал получен при изучении сдвигов ПП на различные афферентные раздражения как при адекватной стимуляции рецепторных образований, так и при электрической стимуляции афферентных путей. Эти данные в значительной степени противоречивы. Противоречия касаются, во-первых, знака сдвигов ПП, а во-вторых, степени распространенности сдвигов по коре мозга, что, возможно, в некоторой степени зависит от применяемой авторами техники отведения и особенно качества и характера электродов. Локальные сдвиги ПП большинство авторов находили в коре кошек и собак, а диффузные сдвиги, у кролика. Однако, как было показано в лаборатории В. С. Русинова (1969) на одном и том же животном (кролик), при внешних раздражениях можно получить различные сдвиги ПП в зависимости от типа отводящих электродов, от их давления на поверхность мозга; эти сдвиги могли быть отрицательными или положительными, локальными или диффузными. Сдвиг ПП коры зависит также от силы внешнего раздражения: при слабой стимуляции преобладают сдвиги ПП с положительным знаком, при усилении раздражения — преобладают отрицательные сдвиги.
Ряд исследователей показал, что негативный сдвиг ПП в коре при афферентных раздражениях связан с импульсацией, поступающей по неспецифическим путям, и тем самым с деполяризацией апикальных дендритов корковых нейронов. Однако это не совпадает с данными других исследователей, которые нашли, что, изменяя только параметры раздражителя, можно получить и негативные и позитивные отклонения ПП.
Пытаясь объяснить эти факты, ряд исследователей выдвинули предположения, что сдвиги ПП поверхности коры имеют сложное происхождение и являются результатом алгебраической суммации многих полей ПП с разными знаками. Тогда с изменением места стимуляции или отведения, а также с изменением параметров раздражения изменяется амплитуда тех или иных полей, а отсюда и направление сдвига ПП (хотя пока и не ясно, что является источником этих полей).
Более или менее однозначные общие заключения относительно сдвигов ПП при раздражении подкорковых структур и при афферентных раздражениях сделать трудно. Можно только отметить (Русинов, 1969), что при раздражении неспецифических ядер наиболее выражены сдвиги ПП в передних областях коры, а при стимуляции специфических ядер — в соответствующих проекционных зонах и вокруг них.
При слабых стимулах сдвиги ПП выражены в специфических и близлежащих зонах с преобладанием позитивных сдвигов, при усилении стимула — преобладают неспецифические ответы с преобладанием негативных сдвигов и особенно большое количество таких сдвигов приурочено к передним областям полушарий.
Во время естественных периодов сна — бодрствования ПП поверхности коры зависит от уровня активности животного, во время сна он сдвигается в сторону позитивности, а пробуждение совпадает со сдвигом ПП в сторону негативности, особенно сильной при движениях животного.
Значительные сдвиги ПП коры наблюдаются при условнорефлекторной деятельности. Как правило, сдвиги ПП появляются в коре в начале выработки условного рефлекса и исчезают при его угашении. Эти сдвиги могут иметь различную полярность. На стадии упрочения рефлекса они либо сохраняются, либо исчезают. Более тщательный анализ сдвигов ПП коры при выработке условного оборонительного рефлекса был проведен в лаборатории В. С. Русинова (1969). Было отмечено, что в процессе выработки условной связи сдвиги ПП изменяются не только по характеру их распределения по коре, но и по направлению.
Какова природа и функциональное значение сдвигов ПП в интегративной деятельности нервной системы? В. С. Русинов (1969) полагает, что сдвиги ПП в коре являются «продуктом» нейронного происхождения, хотя пока нет убедительных экспериментальных данных, свидетельствующих, что эти сдвиги действительно связаны с изменениями мембранных потенциалов нейронов.
По предположению Голдринга и О’Лири (Goldring, O’Leary, 1954), медленный потенциал коры при раздражении средней линии таламуса является результатом суммации эффектов последействия от вызванных ответов (при низкой частоте раздражения) или суммации вызванных ответов и эффектов последействия (при высокой Частоте раздражения). Результат суммации рассматривается ими как стойкое состояние активности дендритов. Еще Клер и Бишоп (Clare, Bishop, 1955) показали, что при частых раздражениях возникновение медленного потенциала коры связано с суммацией дендритных потенциалов.
Брукхарт и сотрудники (Brookhart et al., 1958) также получили косвенные данные, что медленный отрицательный потенциал коры при высоких частотах раздражения специфических и неспецифических ядер таламуса зависит от постсинаптической активности корковых нейронов, вызванной приходом таламо-кортикальных импульсов.
Таким образом, было выдвинуто предположение, что сдвиги ПП в коре обусловлены транссинаптической активацией дендритов и суммацией их ПСП.
Однако, по данным В. С. Русинова и сотрудников, ПП не являются продуктом слияния и совпадения отдельных волн возбуждения, а есть отражение местной длительной активности (Русинов, 1969). Стационарный местный потенциал возрастает и падает под влиянием раздражения. По его мнению, медленные сдвиги-ПП отражают стационарное возбуждение. Подобно тому как быстрые изменения потенциала отражают бегущую волну возбуждения, длительные — отражают медленные, долго длящиеся изменения возбуждения. Таким образом, по В. С. Русинову, ПП и его сдвиги отражают активное физиологическое состояние.
Ряд исследователей, как мы уже отмечали, связывают ПП и его сдвиги с глиальными элементами или с нейронно-глиальным комплексом. Это предположение, хотя и вполне правомерно, т. е. имеется ряд экспериментальных данных в его пользу, не может считаться окончательным, так как отсутствуют четкие результаты о функциональных особенностях глиальных клеток коры.
Достаточно спорен еще вопрос о функциональном значении ПП и его сдвигов. Еще Жерар (Gerard, 1936) предположил, что медленно меняющееся электрическое поле в коре может быть фактором, влияющим на нейроны и вынуждающим их работать синхронно.
О влиянии внешнего поля на активность нервных клеток сейчас имеется достаточно данных на самых различных объектах исследования. Однако работ, специально посвященных активности отдельных нейронов на фоне длительно протекающих сдвигов ПП, еще далеко не достаточно.
Известно, что активность корковых нейронов меняется при сдвигах ПП, вызванных искусственной поляризацией коры (Морелл, 1962; Creutzfeldt, 1962; Воронин, Скребицкий, 1964, и др.). Однако ряд авторов, изучая сдвиги ПП при других воздействиях, отрицают их регулирующее влияние на активность кортикальных нейронов (Li, Salmoiraghi, 1963, и др.).
Поскольку в коре мозга нервные элементы тесно прилегают друг к Другу, создается, видимо, реальная возможность для влияния на соседние элементы электрического поля, возникающего в группе сходно ориентированных элементов. Вопрос о возможности взаимодействия между нейронами посредством внеклеточного электрического поля явился предметом специального изучения в лаборатории В. С. Русинова на примере нейрона рецептора растяжения рака (Эзрохи, 1967). Результаты опытов Эзрохи указывают на реальную возможность осуществления внеклеточным электрическим полем, созданным нейронами физиологически значимого эффекта, который должен заключаться в синхронизации активности.
Чувствительность нейронов ЦНС к модуляции их спайкового разряда под влиянием внешнего поля может быть, видимо, даже выше, чем у нейронов рецептора растяжения рака (Strumwasser, Rosenthal, 1960). Наличие влияния электрического поля одного нейрона на другой показано для эпилептического очага. В этом случае в нейронах наблюдаются пароксизмальные деполяризационные сдвиги и регистрируются высокоамплитудные
внеклеточные потенциалы, которые могут, видимо, служить стимулом для инициации клеточных разрядов в сома-дендритной мембране соседних нейронов, что способствует синхронизации последних (Беленков, Чирков, 1961, 1965; Беленков, 1967 и др.).
Эти экспериментальные данные о регулирующей, роли внеклеточных полей нейронов хорошо подкрепляют гипотезу о ведущей роли медленно изменяющихся потенциалов в функции замыкания временных связей, выдвинутой в 1953 г. В, С. Русиновым. Динамика сдвигов ПП при выработке временных связей, их устойчивость при упрочении связи, исчезновение при угашении и выработке дифференцировки свидетельствуют, по его мнению (1969), в пользу предположения «о ведущей роли стационарных полей», т. е. медленно изменяющихся потенциалов в функции замыкания временной связи
Таким образом, изучение генеза ПП коры и его сдвигов при различных воздействиях показывает, что они играют, видимо, определенную роль в интегративных корковых функциях, исследование которых по сути дела, только начато.
Удельный вес ПП коры и его изменений в генезе суммарной электрической активности мозга, видимо, значителен, так как целый ряд ЭЭГ феноменов, связанных с сигнальной деятельностью у животных и человека, с важными функциональными сдвигами (сон — бодрствование) и рядом патологических изменений ЭЭГ сопровождается, как правило, и изменением ПП. Причинную связь этих процессов еще необходимо исследовать.
Сверхмедленные ритмические колебания в коре больших полушарий
Сверхмедленные (или «инфрамедленные») ритмические колебания потенциалов в коре больших полушарий были открыты, описаны и изучены Н. А. Аладжаловой и сотрудниками (1956, 1957, 1962 и др.). Они свойственны как мозгу животных, так и человека (Аладжалова, 1962; Norton, Jewett, 1965). Общую их характеристику мы дали ранее.
➥ Более подробно: Общая классификация различных видов электрической активности головного мозга и их краткая характеристика
Сверхмедленные колебания потенциала (СМКП) отличаются от апериодических сдвигов ПП тем, что являются ритмическими незатухающими колебаниями, регистрирующимися в течение многих часов и суток. Н. А. Аладжалова (1962) полагает, что СМКП возникают в иной электрохимической системе, чем сдвиги ПП мозга.
СМКП могут быть зарегистрированы во всех зонах полушария и могут появляться не одновременно, причем частота и амплитуда их в разных зонах коры также может быть разной. Они могут быть синхронными в разных точках полушарий (особенно при действии внешних раздражений) и несинхронными. Как правило, они регистрируются у бодрствующего животного, причем их регулярность значительно возрастает при систематическом влиянии на организм, в том числе и при изменении метаболизма нервной ткани.
Так, усиление СМКП наблюдается при длительных и повторных раздражениях рецепторов, при введении гормонов, фармакологических веществ, стимуляции гипоталамической области мозга, т. е. при воздействии на разные звенья метаболизма. В головном мозге человека СМКП особенно усиливаются при «стресс-состоянии», что, по мнению Н. А. Аладжаловой (1969), также указывает на их связь с нейроэндокринными факторами.
Н. А. Аладжалова считает, что для СМКП существует специальная управляющая (или регулирующая) система, в отличие от известных регулирующих систем для более быстрых ритмов и их изменений.
Так, аппарат усиления СМКП включается только тогда, когда внешний стимул действует систематически. Это побуждает медленную управляющую систему перестраивать деятельность мозга на новый уровень. Так, например, однократная стимуляция ретикулярной формации сразу вызывает реакцию активации ЭЭГ, по не влияет на СМКП. Усиление СМКП возникает лишь через 30—40 мин при многократном раздражении гипоталамуса.
➥ Более подробно: Некоторые общие механизмы регуляции спонтанной электрической активности головного мозга

А — электрограммы, расположенные подряд; Б — сверхмедленные колебания потенциала, зарегист-рированные вне раздражения (по Аладжаловой, 1969)
Стрелки указывают момент стимуляции полоски в течение 5 с импульсами 2 мс и 15 Гц (активность возрастает через 1 мин после стимуляции и длится 2—3 мии). Площадь полоски 4X6 см², толщина 180 мкм
Самостоятельное существование сверхмедленной активности (независимо от быстрой) Н. А. Аладжаловой (1962) удалось показать на изолированной от нервных связей полоске коры. В такой деафферентированной полоске коры при состоянии покоя обычная синаптически вызванная активность резко угнетается или исчезает, в то время как СМКП остаются и их параметры могут быть изменены внешними воздействиями (рис. 3).
Такое усиление СМКП в полоске коры может происходить лишь в результате действия химических факторов, поступающих через внутреннюю среду. Предполагается, например, что ингибитор холинэстеразы является одним из эффективных веществ, усиливающих СМКП. Эффект его действия после введения в вену двойной: усиление СМКП наблюдается дважды — в течение первых минут после введения (непосредственное действие на тканевую холинэстеразу) и в период 60—200 мин после введения (результат изменений гормональных взаимоотношений, опосредованные через включение в реакцию гипоталамической области) (Аладжалова, 1962). Подтверждением последнего является усиление СМКП в полоске коры при электрическом раздражении преоптических ядер гипоталамуса.
Эти, а также ряд других данных, полученных Н. А. Аладжаловой и сотрудниками свидетельствуют о правильности представления, о наличии в мозге медленной управляющей системы, имеющей химическое гуморальное звено, которое включается через значительное время после того или иного воздействия.
В 1962 г. вышеназванный автор выдвинула предположение, что СМКП либо возникают на мембране дендритов, либо вначале в глиальных элементах, а деполяризация дендритов наступает вторично. Предполагалось, что СМКП являются электрическим проявлением очень медленных процессов изменения мембранного потенциала, в происхождении которых особую роль играют биохимические, гормональные факторы. С другой стороны, электрическое поле СМКП должно быть фактором, вовлекающим соседние нейроны в единый ритм изменений. Предполагалось, что внеклеточные токи, созданные источниками СМКП в дендритах нейронов, могут вызывать периодическую де- и гиперполяризацию мембраны дендритов соседнего нейрона. Источниками активных веществ, способных вызвать в дендритах появление СМКП, могут быть как синаптическая, так и тканевая продукция гормонов; причем в последней необходимо учитывать роль глии (Аладжалова, 1962).
В последующие годы Н. А. Аладжалова (1969), опираясь на собственные материалы исследований и литературные данные, приходит к выводу, что большую и, возможно, определяющую роль в происхождении СМКП играют процессы, развивающиеся на границах раздела между нервными и глиальными элементами. А сама сверхмедленная активность связывается с активностью аксонов и дендритов нейронов, а также отростков глиальных клеток. Тела нейронов не участвуют в этом процессе.
В основу этих предположений легли следующие факты.

Биполярное отведение под диплацином (по Аладжалового 1969)
Во-первых, для возникновения СМКП не обязательно наличие тел нейронов. Так, СМКП зарегистрированы в препарате «аксо-дендритная полоска» (Аладжалова, Коштоянц, 1962), которая состояла из двух верхних слоев коры мозга кролика и содержала только дендриты и аксоны нервных элементов и отростки глиальных клеток. Кроме того, СМКП зарегистрированы в специфических глиальных клеточных ансамблях (Brazzie, Neal, 1954), в области area postrema (рис. 4). Во-вторых, было показано (Аладжалова, 1965), что электрическое сопротивление этой области продолжает изменяться в течение получаса после одноминутного воздействия и поэтому не исключено, что многоминутные следовые эффекты, которые развиваются после кратковременного воздействия в разных образованиях мозга, могут быть опосредованы через вовлечение в реакцию нейроглии.
И, наконец, известно, что усиление СМКП в нервной ткани тесно связано с метаболическим напряжением. Сдвиги энергетического обмена могут определяться, например, накоплением К+ в нейроглиальных щелях, высокой активностью АТФ-азы и т. д. Известно (Orkandetal. 1966), что во время импульсной активности нервных волокон зрительного нерва в экстраклеточной среде накапливается К+, который деполяризует мембрану глиальной клетки.
Эти, а также ряд других фактов позволили Н. А. Аладжаловой (1969) представить образование секундного СМКП в виде определенного цикла: активность нервного элемента→накопление К+ в экстраклеточной среде→рост порога нервного элемента→активация АТФ-азы→деполяризация глии→исчезновение К+ в экстраклеточной среде→уменьшение порога нервного элемента. Этот цикл занимает несколько секунд.
Так как СМКП является автоколебательным процессом, то необходим источник энергии для этого процесса, который должен возбуждаться в какой-то момент колебаний. Автор полагает, что источником может быть энергия фосфатного обмена, причем звено активации нервного элемента может и полностью отсутствовать (генерация СМКП в чисто глиальных образованиях). Инициаторами таких циклов, видимо, являются биохимические и физико-химические сдвиги, непосредственно возбуждающие глиальные клетки.
Естественно встает вопрос, какова же роль СМКП, если они могут протекать независимо от нейронного звена?
Н. А. Аладжаловой и О. X. Коштоянцем (1967) было показано, что у большинства нейронов их ритм не подчиняется ритму СМКП. Ритм нейронов подчиняется закону детерминированного взаимодействия между самими же нейронами. Иными словами, «нейроны связаны по закону сильного взаимодействия, а СМКП отражают динамику слабого взаимодействия нейронов с окружающей их средой»
Однако предполагается, что при ряде воздействий и изменении ряда условий, когда резко усиливается влияние среды, СМКП возрастают по амплитуде и начинают влиять на условия взаимодействия между нейронами, изменяя связи между ними, например, путем блокады синапсов, изменения их порогов и т. д.
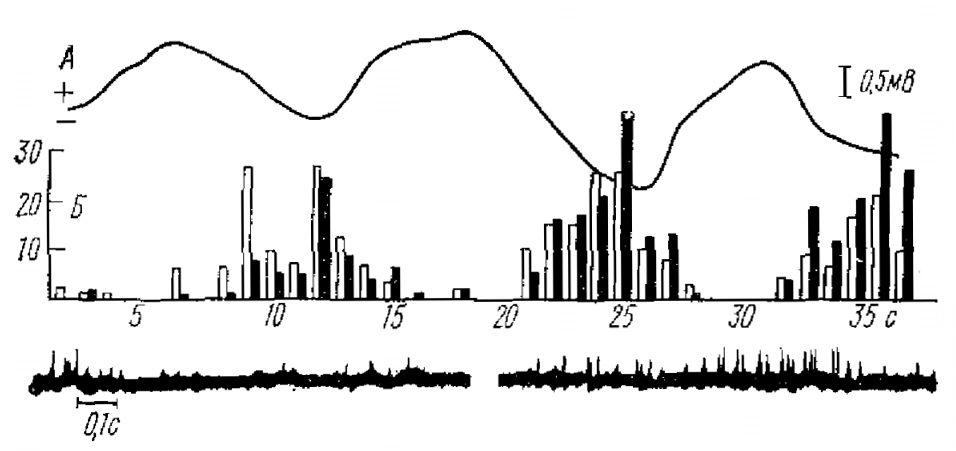
По оси ординат — число разрядов за 1 с, по оси абцисс — время (с), соответствующее шкале времени для сверхмедленного потенциала. Под графиком— записи активности нейронов на положительном и отрицательном гребнях сверхмедленной волны. Монололярная запись на бодрствующем кролике с глубины 1,25 мм
В момент появления высокоамплитудных и регулярных СМКП изменяется и форма ЭЭГ, видимо, за счет образования групповых объединений нейронов или других процессов (Аладжалова, 1969). Экспериментальные данные показывают (Аладжалова, 1969; Fromm, Bond, 1964, и др.), что процент нейронов коры, разряжающихся в такт с СМКП, не очень велик и не удается установить определенной корреляции между фазой, например, секундных СМКП и частотой спайковой активности нейронов, хотя определенные «спонтанные» вспышки активности нейронов, видимо, могут иногда совпадать с секундным и минутным ритмом СМКП (рис. 5). Этот вопрос пока очень слабо исследован. Что касается медленной суммарной активности коры, то она в некоторых случаях закономерно изменяется в такт с ритмом СМКП. Так, степень синхронизации колебаний в ЭЭГ несколько меняется на максимуме и на минимуме волны СМКП. Во время судорожной активности периодическая смена серий колебаний с разной частотой также совпадает с переходом одной фазы СМКП в другую. При ряде воздействий (фармакологических воздействий, раздражении ретикулярной формации, гипоталамуса и т. д.) связь между ЭКоГ и фазой СМКП выступает наиболее явно (Аладжалова, 1969).
Данные об изменении в ряде случаев характера электрической активности коры на максимумах СМКП говорят, видимо, о связи между синаптической активностью нейронов и СМКП. По данным Н. А. Аладжаловой (1969), наибольшую связь с фазой СМКП выявляют изменения активности дендритов корковых нейронов. Таким образом, по ее мнению, взаимодействие между элементами медленной управляющей системы мозга (СМКП) и быстродействующей (ЭЭГ) на уровне коры мозга осуществляется путем влияния тех элементов коры, которым свойственны СМКП, на возбудимость других элементов, функция которых связана только с быстрыми формами активности. Говоря иными словами, ЭЭГ и СМКП обусловлены двумя разными системами генерации суммарной электрической активности мозга разного частотного диапазона, различными по своему генезу. Однако они могут находиться в прямом взаимодействии, механизм и функциональное значение которого еще далеко не ясны.
Она нашла, что структуры головного мозга по признаку выраженности в них СМКП можно разделить на две группы: в одних отсутствуют (ретикулярная формация ствола, таламус, центральное серое вещество), в других они хорошо выражены (область гипоталамуса). В коре головного мозга имеются также две категории элементов: в одних возможно проявление СМКП, в других нет.
Однако электрическая и фармакологическая стимуляция некоторых подкорковых образований, особенно гипоталамуса, может вызвать СМКП в РФ, таламических ядрах, центральном сером веществе и коре. Факторы такого усиления могут быть, видимо, весьма разнообразными, Особую роль придают гуморальным влияниям.
Анализ сроков усиления СМКП в подкорковых образованиях мозга при таких воздействиях позволил Н. А. Аладжаловой (1969) выделить разные звенья, которые обладают следующими тремя признаками.
- Не отвечают на кратковременные и случайные факторы, а включаются при систематических или чрезмерных воздействиях.
- Латентный период их реакций может исчисляться десятками минут.
- Участвуют в регулировании в течение нескольких часов. Эти признаки она относит к свойствам медленной управляющей системы мозга.
Медленный отрицательный потенциал поверхности коры
Одиночное электрическое раздражение коры пороговым стимулом вызывает дендритный Потенциал длительностью 20—30 мс в виде одиночного отрицательного колебания. Он отражает ВПСП верхушечных дендритов (Ройтбак, 1969).
При усилении раздражения после пего развивается медленный отрицательный потенциал (МОП) большой длительности (Ройтбак, 1963; Chang, 1954, и др.). Он был подробно описан и исследован Ройтбаком (1965, 1969).
Длительность МОП достигает 3000 мс, причем с усилением раздражения она значительно возрастает.
Изучение электрического поля (Ройтбак, 1969) показало, что во время МОП возникают радиально ориентированные диполи через всю толщину коры.
Ройтбак (1965, 1969) пришел к выводу об участии нейроглии в происхождении МОП в связи с накоплением в глио-нейрональных щелях химического вещества, очевидно ионов К+, вызывающего деполяризацию верхушечных дендритов.
➥ Более подробно: Гипотезы о природе суммарной электрической активности головного мозга
Косвенным подтверждением такого взгляда на происхождение МОП явились данные Караяши и Голдринга (Karahashi, Goldring, 1966) о внутриклеточном изучении некоторых клеток коры, обычно не дающих ни разряда повреждения, ни отвечающих на раздражения импульсами и обладающих высоким, до 90 мВ мембранным потенциалом. Пороги вызова деполяризации этих клеток коры совпадали с порогом вызова МОП, причем деполяризация их мембраны градуально увеличивалась при усилении интенсивности раздражения и наблюдалась суммация ответов. Деполяризация мембраны этих клеток соответствовала по характеру и длительности МОП на поверхности коры. Логично было связать деполяризацию этих молчащих клеток коры с происхождением МОП, хотя доказательств, что это именно глиальные клетки, у авторов не было. Кроме того, экспериментальный материал свидетельствовал о том, что сам МОП отражает деполяризационные процессы, имеющие место и в апикальных дендритах корковых нейронов. Значит, принимая во внимание глио-нейрональную гипотезу происхождения МОП, необходимо решить вопрос о механизмах глио-дендритных взаимоотношений. Кроме того, далеко еще не был решен вопрос, каким образом активируется сама нейроглия коры больших полушарий. Существенным подтверждением глио-нейрональной природы МОП явилось его избирательное устранение при действии даже малых доз рентгеновских лучей (Ройтбак, 1969) (без заметного изменения дендритного потенциала); в то же время есть данные о высокой чувствительности к ним нейроглии. Однако в 1973 г. А. И. Ройтбак и сотрудники анализируя одновременную запись микроэлектродом от глиальных клеток коры и суммарную запись активности с поверхности коры, приходят к заключению о глиальном происхождении МОП, полагая, что нервные импульсы вызывают деполяризацию глии за счет выделения К+ из возбужденных нервных элементов и за счет выделения медиатора из окончаний нервных волокон.
Есть и другие точки зрения на происхождение МОП. Одна из них, например, связана с представлением о том, что МОП отражает деполяризацию афферентов коры, что вряд ли обоснованно, так как в хронически изолированной полоске коры, где афферентные волокна пере-рождаются, МОП сохраняется.
Вторая точка зрения связывает МОП с гиперполяризацией глубоких частей пирамидных нейронов (Li, Chou, 1962). В этом случае отрицательность МОП на поверхности коры является лишь дипольным отражением ТПСП на глубоких частях пирамидных нейронов.
Однако надо полагать, что эти явления имеют различный генез, тем более что Ройтбак (1973) показал, что МОП отражает в основном активность элементов верхних слоев коры и не является отражением гиперполяризации нейронов глубоких ее слоев.