После открытия и первого подробного описания спонтанной электрической активности коры больших полушарий человека и животных долгое время не предпринимались попытки экспериментально обосновать ее природу. Вначале считалось несомненным, что сложная электрическая активность ЦНС определяется такими же биоэлектрическими процессами, как и в периферических нервах. Так, например, классическим представлением было положение, выдвинутое еще в 1925 г. Шеррингтоном, что центральное возбуждение аналогично местному возбуждению в нерве.
Интересно отметить, что в 1931 г. Эдриан и Байтендайк, изучая ритмическую активность (дыхательные медленные волны) в стволе мозга золотого карася, предположили, что эти потенциалы отражают медленные деполяризационные изменения сомы и дендритов нейронов. Однако позже, в 1934 г., такое предположение о природе медленных колебаний было оставлено самим же Эдрианом. В совместной работе с Мэттьюзом (Adrian, Mallows, 1934) и последующих его работах были сформулированы основные выводы о происхождении медленных электрических колебании головного мозга, которые надолго определили подходы и методы экспериментальных исследований в области электрофизиологии центральной нервной системы.
Содержание этой гипотезы определялось наиболее исследованной в то время формой элементарной нервной активности в виде распространяющегося пикового потенциала действия нервного проводника.
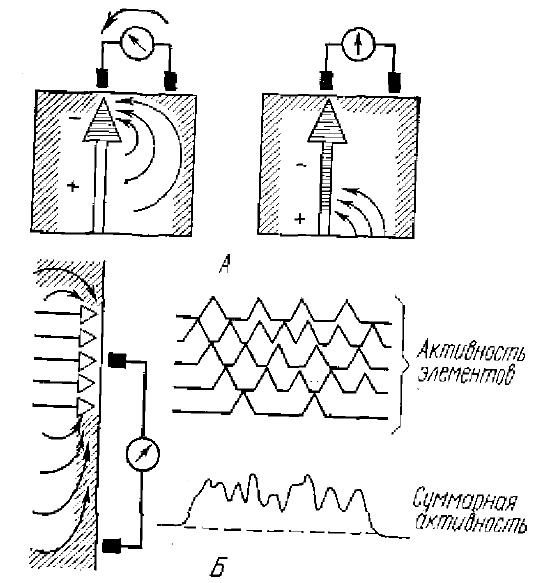
А — токи, возникающие при появлении в нервной клетке волны распространяющегося возбуждения, движущейся вниз по аксону, от поверхности коры при этом должна регистрироваться монофазная волна; Б — токи и потенциалы, возникающие при ритмической асинхронной активности группы нервных клеток; суммарная активность формируется за счет суммации монофазных потенциалов действия
Эдриан и Мэттьюз предположили, что медленные потенциалы коры представляют собой суммацию монофазных пиковых потенциалов длительностью от 10 до 100 мс, возникающих не в аксоне корковых нейронов, а в их соме или дендритах. Таким образом, ими постулировалось, что длительность липовых потенциалов сомы и дендритов должна превышать 10 мс. По их теории каждый импульсный разряд одиночного нейрона создает на поверхности коры небольшое монофазное колебание потенциала. Одновременное их возникновение и частичное перекрытие во времени во многих корковых нейронах приводит к значительному суммарному колебанию потенциала, которое можно зарегистрировать (рис. 1).
Подобное явление было известно в то время для целого нерва, т. е. было показано, что суммарные медленные его потенциалы складываются из потенциалов действия отдельных нервных волокон. Было также известно, что частичная дисперсия во времени потенциалов действия отдельных нервных волокон приводит к удлинению суммарного медленного потенциала действия целого нервного ствола.
Таким образом, предполагалось, что значительная длительность суммарных электрических колебаний коры является также следствием временной дисперсии составляющих их потенциалов — потенциалов действия нервных клеток. Колебательный характер суммарной электрической активности в этом случае обусловлен периодическими изменениями частоты импульсной активности. При относительно синхронном увеличении частоты импульсной активности в каждый момент времени возникают импульсные разряды в большом количестве нервных клеток, результатом чего явится регистрация значительного потенциала на поверхности коры. Затем следует уменьшение общей частоты импульсной активности нейронов, поэтому в каждый момент времени суммируются импульсы от меньшего количества нейронов, — величина суммарного колебания падает. Периодическое повторение этих процессов создает картину ЭЭГ, регистрируемую с поверхности коры, где периодичность и амплитуда волн определяется характером периодических изменений частоты импульсной активности в относительно большой популяции нейронов. Естественно, от этого зависит и рисунок кривой суммарной ЭЭГ, Существенным в гипотезе было и то, что периодические изменения частоты активности должны происходить синхронно в большом количестве нейронов, что необходимо для возникновения суммарных электрических колебаний.
Сейчас гипотеза Эдриана и Мэттьюза имеет лишь исторический интерес.
Начиная с 40-х годов накапливаются новые данные, противоречащие теории Эдриана и Мэттьюза. Уже первые микроэлектродные исследования Реншоу и сотрудников (Renshaw et al., 1940) на отдельных нейронах коры и гиппокампа поставили под сомнение природу ЭЭГ как сумму потенциалов действия. Трудно было предположить, чтобы потенциалы действия нейронов, равные в среднем 1,5 мс (а не выше 10 мс, как полагал Эдриан), могли суммироваться и создавать волны ЭЭГ продолжительностью от 10 до 1000 мс. Наиболее веские доказательства были получены значительно позже в опытах на животных при одновременной регистрации медленной и импульсной активности. В работах Ли и соавторов (Li et al., 1952; Li, Jasper, 1953) было показано, что импульсная активность нейронов коры мозга значительно более чувствительна к ухудшению функционального состояния, чем медленная суммарная электрическая активность. Легкая аноксия или глубокий наркоз полностью подавляли импульсные разряды, тогда как суммарные электрические колебания сохранялись и были хорошо выражены. Не было показано и определенного соответствия между спайковой активностью и медленными волнами ЭЭГ. Наблюдалась даже обратная картина; в одном и том же участке коры активность некоторых нейронов угнеталась во время вспышки медленных волн в ЭЭГ.
Соотношение суммарной медленной и нейронной активности изучалось в работе Бухвальда и сотрудников (Buchwaid et al., 1956). Ими была использована специальная методика одновременной регистрации одним электродом медленной суммарной активности (ЭЭГ) и активности многих нейронов. Было показано, что возможны три варианта, когда: 1) активность нейронов меняется, но без всяких соответствующих изменений суммарной ЭЭГ; 2) ЭЭГ испытывает значительные изменения без каких-либо изменений в активности нейронов, и 3) изменения нейрональной активности совпадали с изменением медленной активности мозга.
Об отсутствии четкой взаимосвязи между спайковой и медленной электрической активностью свидетельствуют и данные других исследователей (Stopp, Whitfield, 1963; Анохин, 1964, Робертсон, 1965, и др.).
Итак, все эти данные не подтверждали вывод о том, что ЭЭГ является результатом спайковых разрядов нейронов и что медленные колебания ЭЭГ и нейрональная активность находятся в прямой зависимости. Можно было сделать заключение, что суммарная активность мозга должна складываться из более длительных, градуальных электрических потенциалов, каковыми могут являться синаптические и особенно дендритные потенциалы, тем более что были получены факты, показывающие, что деполяризационные потенциалы корковых нейронов даже при значительной амплитуде их колебании не обязательно приводят к генерации спайковых потенциалов. Отсутствие корреляции (или ее наличие) между медленной активностью коры и спайковыми разрядами нейронов можно удовлетворительно объяснить на основе гипотезы, согласно которой медленные потенциалы, регистрируемые в поверхностных (пли глубоких) слоях коры, состоят из деполяризационных или гиперполяризационных постсипаптических потенциалов, генерируемых в сложноорганизованных элементах — соме и дендритах корковых нейронов.
В этом отношении интересно сравнить данные Клее с соавторами (Klee et al., 1965) и Крейцфельда с соавторами (Creutzfeldt et аl.. 1966). Если первые авторы показали, что коэффициент кросс-корреляции между ЭЭГ волнами и колебаниями мембранного потенциала нейронов достигает величины более 07, то вторые нашли, что максимальная корреляция между ЭЭГ и клеточной импульсной активностью колеблется в пределах только 0,4—0,7.
Однако вопрос о том, какие медленные потенциалы лежат в основе тех или иных феноменов ЭЭГ, до сих пор полностью не решен. Это связано, видимо, с тем, что для коры и других мозговых образований характерно наличие большого спектра медленных и сверхмедленных колебаний потенциалов, в происхождении многих из которых еще в достаточной степени гипотетично и в основе которых могут лежать самые разные по происхождению колебания, генерируемые теми или иными функциональными образованиями нейронов или другими клеточными элементами коры больших полушарий. То же относится, видимо, и к разным видам суммарной активности других нервных образований, подкорки, мозжечка и т. д.
Таким образом, все последующие работы вплоть до 70-х годов убедительно показали неприемлемость гипотезы Эдриана и Мэттьюза к корковому электрогенезу. Трудно определить, когда потенциалы коры стали считаться характеристикой клеточных, медленных потенциалов. Безусловно, появлению новых гипотез об электрогенезе мозговых волн способствовал ряд новых данных, полученных в 30—40-е годы.
Первым фактором явилось открытие способности нейронов коры генерировать медленные волны, вторым — признание особых свойств дендритов, и, наконец, третьим— открытие того, что медленные потенциалы связаны с синаптической передачей. Именно в свете этих данных и была выдвинута новая гипотеза об электрогенезе суммарной электрической активности коры, которая опиралась на принципы теории синаптического электрогенеза и известные свойства дендритного потенциала. Вначале эта гипотеза встречала определенные трудности, так как была еще далеко не ясна природа медленных потенциалов нейронов, которую объясняли либо активностью пресинаптических окончаний, либо постсинаптических элементов или же тех и других вместе. Открытие медленных синаптических возбуждающих и тормозных потенциалов и первые данные о свойствах дендритов определило весь дальнейший ход рассуждений.
Была выдвинута более обоснованная гипотеза, когда суммарная электрическая активность коры стала рассматриваться как результат суммации в основном постсинаптических градуальных потенциалов корковых нейронов, где наиболее важную роль в ее электрогенезе начинают усматривать в особых свойствах дендритов корковых нейронов.
Еще в 1943 г. О’Лири и Бишоп (O’Leary, Bishop) высказались за то, что медленная суммарная активность является дендритной активностью.
В последующих работах Чанга, Тасаки, Бишопа, Пурпуры, Грундфеста, Ройтбака и др. исследователей изучаются свойства дендритов, которые приводят к выводу, что суммарная медленная активность возникает в обширной сети дендритных ветвлений. Этот вывод подкреплялся многими фактами: совпадением онтогенетического формирования активности коры и созреванием ее апикальных дендритов; способностью дендритов к суммации (а отсюда—длительность дендритных потенциалов, перекрывающая по временным характеристикам все виды медленной суммарной активности мозга); наличием широких дендритных ветвлений, составляющих основную массу коры и хорошо выраженных медленных колебаний в области их ветвлений; наличием однотипного усиления суммарной активности и прямого (дендритного) ответа коры под действием анода постоянного тока и идентичного их угнетения при действии катода и т. д.
Однако, как показали экспериментальные данные, в ЭЭГ, регистрируемой с поверхности коры, должны отражаться не только ПСП апикальных дендритов, но и ПСП глубоких частей пирамидных нейронов — сомы и базальных дендритов.
Необходимо отметить, что гипотезе электрогенеза ЭЭГ как суммы постсинаптических градуальных потенциалов отнюдь не противоречат данные о наличии при определенных условиях (спонтанная или вызванная ритмическая активность мозга, прямое раздражение коры, эпилептическая активность, фармакологические воздействия и т. д.) четкой корреляции между спайковой и медленной активностью. Механизмы такой четкой корреляции будут разобраны нами в последующих лекциях (см. лекции по генезу альфа-активности, бета-ритму, реакциям вовлечения и усиления и др.).
Исследования последних лет в значительной степени детализировали и дополнили теорию постсинаптической сома-дендритной природы суммарной медленной активности мозга, которая до сих пор является основной теорией.
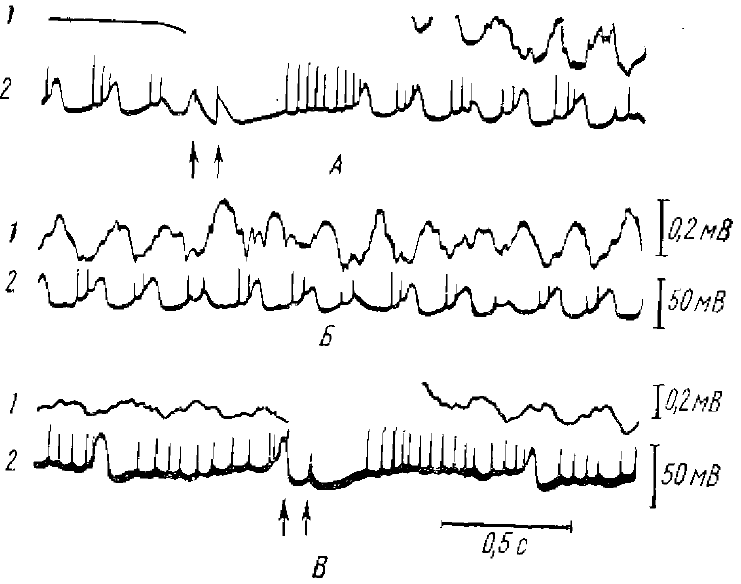
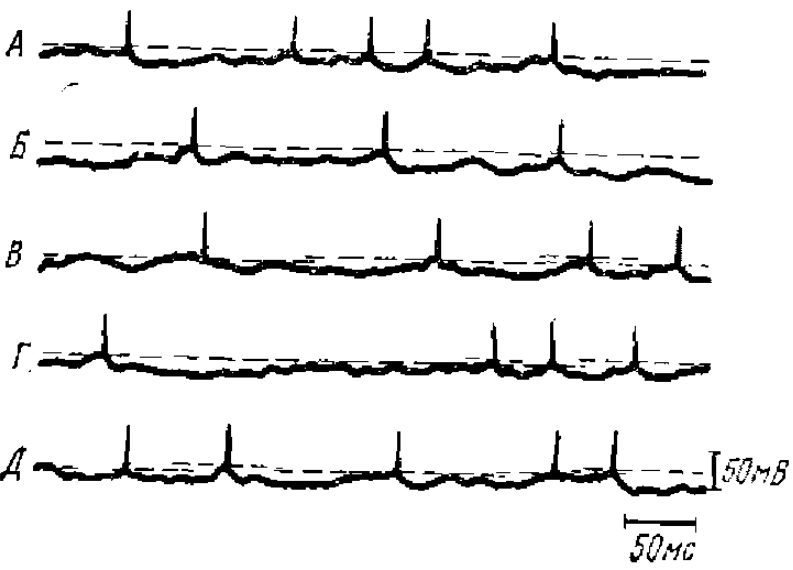
Стрелками отмечено время раздражения, вызывающего θ -ритм. Для суммарной и внутриклеточной регистрации отклонение вверх соответствует позитивности активного электрода
Внутриклеточные исследования многих авторов показали, что существует тесная связь между синаптической активностью отдельных нейронов и поверхностно peгистрируемой ЭЭГ. Так, сопоставление внутриклеточной активности гиппокампа с суммарной его активностью (Fujita, Sato, 1964) показало, что мембранный потенциал изменяется синхронно с суммарной активностью (рис. 2). Близкие данные на новой коре были получены Калветом с соавторами (Calvet et al., 1964).
Детальное исследование внутриклеточных колебаний потенциала в связи с суммарными корковыми потенциалами спонтанной веретенообразной активности было проведено Джаспером и Стефанисом (Jasper, Stcfanis. 1965) на нейронах пирамидного тракта кошки.
Веретенообразная активность, регистрируемая от поверхности коры, состояла преимущественно из негативных колебаний. Одновременная регистрация поверхностной электрокортикограммы и внутриклеточных потенциалов показала, что между ними наблюдается хорошее соответствие. Из этих данных было сделано заключение, что основную роль в генерации корковых потенциалов спонтанной веретенообразной активности играют деполяризующие синапсы, расположенные в поверхностных слоях коры.
Элул (Elul, 1964, 1969) провел серию исследовании по соотношению медленной активности коры и внутриклеточной активности ее нейронов и обнаружил у ненаркотизированных животных в корковых нейронах большие ритмические волны с амплитудой от 5 до 15 мВ, что составляло значительную часть мембранного потенциала нейрона, который колебался от 70 до 90 мВ, и превышало в сотни раз амплитуду суммарной ЭЭГ, записанной от прилежащей внеклеточной ткани и с поверхности коры. Между колебаниями внутриклеточной активности и суммарной ЭЭГ наблюдалось (хотя и кратковременное) четкое соответствие (см. рис. 7).
Эти и другие исследования явились веским подтверждением гипотезы о природе ЭЭГ как суммации ПСП корковых нейронов.
Некоторое разногласие между исследователями вызывала первое время трактовка расхождения по фазе регистрируемых внутриклеточно ПСП и волн ЭЭГ, регистрируемых с поверхности коры. Одни авторы полагали, что возбуждающие ПСП связаны с поверхностно положительными волнами ЭЭГ, а другие эти волны связывали с тормозными ПСП. Те же разногласия касались и поверхностно-отрицательных волн ЭЭГ. Как показал дальнейший анализ, необходимо было учитывать явление диполя, возникающего в нейронах коры, имеющих направленные апикальные дендриты (рис. 3). Благодаря такому расположению значительной массы корковых нейронов при интерпретации медленных волн ЭЭГ, регистрируемых с поверхности коры, необходим в каждом случае специальный анализ тех синаптических процессов, отражением которых они являются.
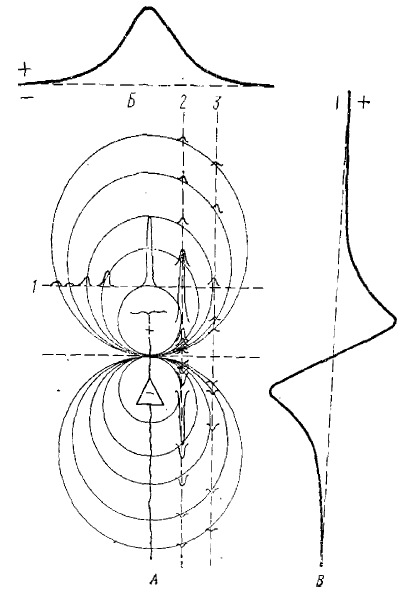
Тонкие пунктирные линии покатывают сечения через это поле, перпендикулярные к оси диполя (1) или параллельные ей (2, 3), Приведена также амплитуда и полярность отклонений в соответствующих точках поля. Б — пространственное распределение потенциала диполя в плоскости (1). В — пространственное распределение потенциала диполя в плоскости (2)
Из сказанного можно сделать вывод, что электроотрицательная волна ЭЭГ, регистрируемая с поверхности коры, может определяться ВПСП верхушечных дендритов или (в результате дипольного отражения) ТПСП,
Так, например, в коре больших полушарий имеется несколько синаптических систем, принимающих участие в генерации веретенообразной активности. Это подтверждается детальными исследованиями распределения по поперечнику коры потенциалов веретенообразной активности (Calcci et al, 1961, 1964). Применение в этих опытах специального отведения потенциалов коры позволило локализовать полюсы генераторов электрической активности в слоях коры. В результате было установлено, что существует три различных генератора веретенообразной активности.
Первый генератор создает разность потенциалов в поверхностных слоях коры (от поверхности до глубины 500 мкм). При этом возникают колебания потенциала, имеющие негативную полярность на поверхности коры. Эта негативность связана, видимо, с системой деполяризующих синапсов в поверхностных слоях коры.
Второй — создает разность потенциалов между поверхностью и глубокими слоями (около 900 мкм) коры; он генерирует колебания, имеющие позитивный знак на поверхности и негативный знак в глубине коры, Этот генератор может быть системой деполяризующих синапсов в глубоких слоях коры (позитивность на поверхности является дипольным отражением негативности глубоких слоев).
Наконец, третий генератор создает разность потенциалов между поверхностью и той же глубиной коры (около 900 мкм), а генерируемые колебания негативны на поверхности коры и позитивны в глубине. Этим генератором, очевидно, являются гиперполяризующие синапсы, расположенные также в глубоких слоях коры. Отсюда ясно, что медленные колебания, регистрируемые от поверхности коры, определяются алгебраической суммой ВПСП и ТПСП, возникающих в разных слоях коры, а характер этих медленных колебаний (амплитуда, длительность, смена фаз и т, д.) зависит от временной последовательности работы этих генераторов и амплитуды деполяризационных или гиперполяризационных колебаний на каждом уровне.
Таким образом, сложная кривая ЭЭГ может отражать сложную динамику деятельности различных генераторов, которая зависит от. динамических соотношений афферентных корковых систем и внутрикорковых процессов.
Так, для некоторых видов медленной активности коры причиной образования положительного диполя у поверхности коры могут быть импульсы из специфических афферентов, вызывающие ВПСП на уровне базальных дендритов нейронов III—IV слоя, а в случае отрицательного диполя у поверхности коры — импульсы из неспецифических афферентов. Таким образом, в ЭЭГ, регистрируемой с поверхности коры, должны отражаться не только ВПСП (возможно, и ТПСП) апикальных дендритов корковых нейронов, но с обратным знаком ВПСП и ТПСП глубоких частей пирамидных нейронов, их сомы и базальных дендритов.
Несколько по-иному трактовал дендритную природу суммарной активности мозга Касперс (Caspers, 1959). Он рассматривал колебания потенциалов коры (ЭЭГ) как амплитудную модуляцию так называемого постоянного потенциала коры.
Было показано, что существует постоянно фоновый градиент потенциала коры, который может меняться при различных воздействиях и изменениях функционального состояния ЦНС.
Касперс и Шульце (Caspers, Schulze, 1959) выяснили, что амплитуда, форма и направление дендритного потенциала коры (дендритный ответ коры на электрическое раздражение) определенным образом зависят от уровня постоянного потенциала поверхности коры. При развитии негативности коры наблюдается уменьшение амплитуды дендритного потенциала. Чем выше негативный уровень коры, тем меньше амплитуда поверхностного дендритного потенциала. Такой эффект наблюдается при непосредственной катодной поляризации поверхности коры. При анодной поляризации наблюдается обратный эффект и амплитуда дендритного ответа коры увеличивается. По мере увеличения негативности коры может наблюдаться почти полное исчезновение дендритного потенциала, и если дальнейшая негативность нарастает, то прямое электрическое раздраженно коры вызывает противоположный эффект — позитивное колебание, которое тем больше, чем выше негативный уровень постоянного потенциала. Зависимость амплитуды и знака поверхностного дендритного потенциала от уровня и величины, сдвига постоянного потенциала коры и привела Касперса к выводу, что поверхностно вызванный дендритный потенциал является амплитудной (локально ограниченной) модуляцией постоянного потенциала.
В исследованиях ряда других авторов было показано, что и характер изменения фоновой ритмики ЭЭГ совершенно сходен с характером изменений дендритного потенциала при одних и тех же сдвигах уровня постоянного потенциала коры. Поскольку в то время признавалось, что фоновая суммарная активность коры является продуктом суммации колебаний только дендритных потенциалов, это дало основание Касперсу сделать предположение, что и прямой — дендритный ответ коры и фоновая ритмика — ЭЭГ представляют собой амплитудную модуляцию постоянного потенциала коры.
По гипотезе Касперса, существует два уровня постоянной негативации коры — индифферентный и нормальный уровни. Индифферентный уровень негативности — это такой стабильный уровень поляризации дендритной мембраны, когда ни прямое раздражение коры, ни приток афферентного потока к дендритным образованиям не могут вызвать изменений этой негативной поляризации. Нормальный уровень лежит ниже индифферентного уровня и соответствует спокойному состоянию, степень негативности коры ниже (уменьшается поляризация дендритов). В этом случае прямое раздражение коры отражается в виде отрицательного колебания, что отражает возрастание негативности в направлении индифферентного уровня. Таким образом, и фоновая ритмика, т. е. ЭЭГ, также является амплитудной модуляцией постоянного потенциала коры в зоне между его нормальным и индифферентным уровнями. Причины, вызывающие эти колебания, могут быть самыми разными: фоновая импульсная бомбардировка, внешние воздействия и т. д.
Гипотеза Касперса оказалась неприменимой к объяснению многих ЭЭГ колебаний, электрогенез которых в настоящее время достоверен, но она обратила особое внимание нейрофизиологов на роль сверхмедленных электрических процессов головного мозга.
Другая гипотеза о происхождении медленной суммарной активности мозга основана на данных по изучению возможной роли глии в электрогенезе медленных колебаний мозговых потенциалов. Хотя в настоящее время происхождение основной, суммарной активности мозга никто и не связывает только с активностью глиальных элементов, однако прямое участие последних в происхождении некоторых электрических колебаний мозга, видимо, имеет место.
Глия, по данным разных исследователей, занимает от 30 до 50% серого вещества мозга. Нейроглиальные элементы ЦНС представлены тремя типами клеток: астроцитами, олигодендроцитами и макроглиоцитами. Соотношение этих трех элементов в различных отделах ЦНС и зонах коры больших полушарий различно. Электронная микроскопия показала, что наблюдается такое тесное расположение глиальных клеток и нейронов, что большая часть поверхности сомы и отростков нервных клеток находится в контакте с глиальными клетками. Поэтому неоднократно высказывались предположения о том, что глия влияет на деятельность нейронов, а также должно иметь место и обратное влияние.
Вопрос о способах возбуждения глиальных клеток на афферентную импульсацию является крайне важным. Так, были найдены синаптические контакты на олигодендроцитах коры, что. свидетельствует о возможности распространения импульсов к глиальным элементам мозга. Олигодендроцитов особенно много в IV и верхних слоях коры с соответствующим большим числом нейронно-глиальных контактов. Электронная микроскопия также показала наличие нейронно-глиальных связей и не только с олигодендроцитами, но и с астроцитами, хотя в области таких контактов не обнаруживаются характерные для синапсов утолщения мембран.
Предположения о том, что глия участвует в происхождении медленных потенциалов ЦНС, высказывались многими исследователями. Еще в 1955 г. эта мысль была высказана Глис (Glees), которая впоследствии была развита и поддержана Галамбосом (Galambos, 1961). Последний полагал, что глия ответственна за многие виды медленных волн ЭЭГ, такие, как α-ритм, медленные колебания в ЭЭГ во время сна, при эпилепсии, при опухолях мозга и т. д. Электрогенез ряда медленных и сверхмедленных колебаний потенциала в коре больших полушарий и других отделах мозга связывают с глиальными элементами Ройтбак (1965), Аладжалова (1969) и некоторые другие авторы.
В 1962 г. Е. Н. Соколов также поддержал гипотезу Галамбоса о роли глии в генерации суммарной активности мозга. Он предположил, что «если на приход импульсов клетки глии отвечают возбуждением, при которых ионы калия выделяются в межклеточную среду, то это должно вести к частичной деполяризации нейронов, распространяющейся в определенном пространстве и сопряженной с повышением их возбудимости…».1 Таким образом, медленная активность мозга, по Соколову, связана с рефлекторно регулируемыми колебаниями ионного
В последние годы появилось значительное количество работ, экспериментально показывающих связь некоторых медленных колебаний мозга с глиальными элементами, Однако, вопрос о том, являются ли эти колебания отражением только активности глии или глионейрональных ансамблей, еще требует решения.
Более того, некоторые исследователи (Vatter, 1972, и др.) пытались обосновать гипотезу, что источниками суммарной ЭЭГ являются главным образом глиальные мембранные потенциалы, а также изменения PH крови, процессы, связанные с диффузией NaCl и разностью потенциалов между сомой и апикальными дендритами.
Особый интерес представляют исследования электрических характеристик глиальных клеток. В культуре ткани, при внутриклеточной регистрации от глиальной клетки (Hild et al., 1959, 1962), было показано, что в ответ на прямое электрическое раздражение глиальная клетка отвечала быстро развивающейся деполяризацией, за которой следовало медленное возвращение к потенциалу покоя. В исследованиях на глиальных клетках позвоночных и беспозвоночных животных, в острых опытах были выяснены некоторые особенности возбуждения глиальных элементов (хотя в ряде работ не было точно установлено, что регистрировались именно глиальные элементы).
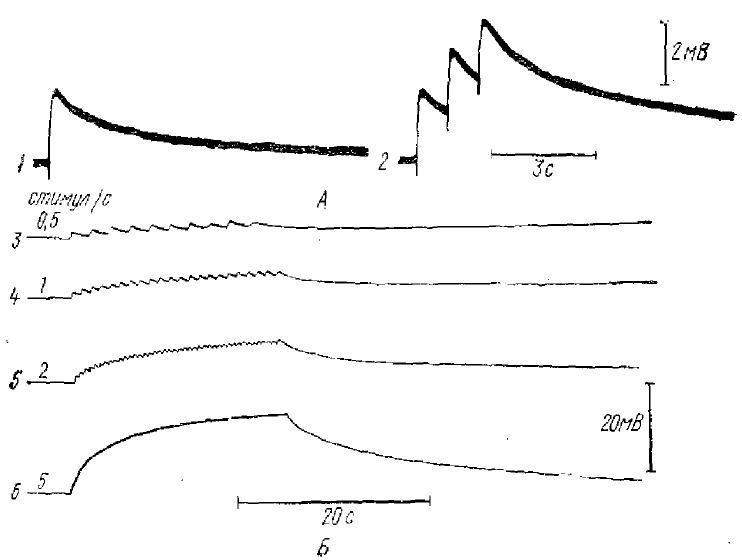
1 — деполяризация в ответ на один стимул максимальной интенсивности, 2 — применение трех стимулов с односекундным интервалом (суммация деполяризации в глиальной клегке), 3—6 — разная частота стимуляции 0,5; 1; 2 и 5 в 1 с
Клетки глии имеют большие потенциалы покоя и не генерируют потенциал действия при деполяризации мембраны: мембранный потенциал может быть смещен в любом направлении до 200 мВ без признаков спайкового ответа. Для большинства глиальных клеток характерна длительная деполяризация мембраны. Так, например, в работах Орканда и др. (Orkand et al., 1966) мембраны глиальных клеток зрительного нерва деполяризовались при раздражении последнего (рис. 4). Ответ достигал максимума за 50—150 мс, а возвращение к уровню покоя длилось несколько секунд. Величина деполяризации достигала 3,1 мВ (рис. 4, А). При ритмических раздражениях происходила суммация ответа. При частоте раздражения 5 в 1 с (в течение 20 с его действия) деполяризация превышала 10 мВ (рис. 4, Б). Деполяризация мембраны глиальной клетки в зрительном нерве, объясняется на основании гипотезы о накоплении калия в межклеточных щелях (за счет импульсной активности нервных волокон зрительного нерва, прилежащих к этой глиальной клетке).
А. И. Ройтбак (1968) считает, что в коре больших полушарий существуют два механизма, посредством которых нервные импульсы могут вызвать деполяризацию мембраны глиальных клеток. Один из них связан с выделением калия из нервных волокон в межклеточные щели; второй — с выделением медиатора из окончаний нервных волокон, имеющих многочисленные контакты с глиальными элементами. Автор считает, что существует и несколько возможных механизмов деполяризации дендритов, приводящей к генерации медленного потенциала коры. В этом случае под действием ионов калия, освобождающегося из возбуждающих нервных волокон, происходит либо деполяризация дендритов и глиальных элементов независимо друг от друга, либо деполяризация глиальных мембран происходит под действием ионов калия и затем глиальные токи разносят их по межклеточным щелям, результатом чего и является деполяризация дендритных ветвлений. Ройтбак также полагает, что деполяризация глиальных мембран связана с действием медиатора; это приводит к выделению ионов калия и последующей деполяризации дендритов.
Однако мы еще очень мало знаем о глионейрональных взаимоотношениях, а также об источниках возбуждения глиальных элементов коры и тем более о «химизме» главного звена — нейроглия-нейрон, определяющего, по мнению многих исследователей, ряд медленных колебаний мозга.
Таким образом, мы кратко рассмотрели некоторые гипотезы о происхождении суммарной активности мозга. Как мы видим, наиболее признанной является гипотеза, рассматривающая медленную активность мозга как алгебраическую сумму ВПСП и ТПСП, возникающих в больших популяциях нейронов головного мозга. Безусловно, нельзя «сбрасывать со счета» активность нервных волокон, глиальных элементов и активную роль метаболических процессов.
Рассмотрев различные представления о природе медленной электрической активности мозга, мы, однако, не решили главного вопроса, какова же природа элементарных генераторов медленной активности головного мозга. Мозг, являющийся объемным проводником, представляет собой проводящую среду, в которой имеет место диффузное распределение огромного количества электрически активных элементов, каковыми являются тела нейронов, аксоны, дендриты и глиальные клетки. Все эти элементы являются источниками внеклеточного тока, создающего потенциальное поле вокруг источника (Жадин, 1967).
Возникает вопрос, является ли нервная клетка одиночным элементарным генератором медленной активности мозга или же она представляет собой ряд независимых друг от друга генераторов (Elul, 1972).
Для решения этого вопроса необходимо вернуться несколько назад. Как мы уже говорили, изучение внутриклеточных регистраций от корковых нейронов дало право предполагать, что между внутриклеточными волнами и спонтанными волнами в ЭЭГ имеется тесная связь. Однако природа этой связи нами не обсуждалась. Здесь могут быть три альтернативы: 1) внутриклеточные волны отражают активность, имеющую место во внеклеточной среде, 2) они являются (как и спайковая активность) действительно источником активности самой клетки, 3) внеклеточные и внутриклеточные процессы вообще между собой не связаны. Таким образом, остается неясным, активно или пассивно возникает волновая активность внутри клетки.
Пассивно возникающая волновая активность внутри клетки может быть продуктом больших внеклеточных электрических полей, вызывающих электрический ток в клетке. И наоборот, волны внутри нервной клетки могут возникать активно как за счет проводимости в синаптических участках на поверхности клетки, так и общих изменений проводимости мембраны.
Исследуя этот вопрос, Элул (Elul, 1972) показал, что в обычных условиях эксперимента волны, регистрируемые внеклеточными электродами, как правило, имеют меньшую амплитуду, чем активность, зарегистрированная тем же электродом, после того как он попадает внутрь клетки. Эти данные исключают возможность пассивного индуцирования внутриклеточных волн внеклеточными полями. Он, а также ряд других исследователей полагают, что вообще в норме, видимо, не существует достаточно больших внеклеточных полей для. образования внутриклеточно регистрируемых в нейроне волн, Что касается внеклеточной волновой активности, то Элул показал, что ее генез прямо определяется внутриклеточной волновой активностью.
Пунктирная линия — уровень генерации спайка
Итак, волновая активность, регистрируемая внутриклеточными электродами, берет свое начало в самих исследуемых нейронах. Здесь подразумеваются две возможности образования волн: либо они образуются в нескольких различных точках на поверхности клетки в результате синаптической активности, либо волнообразное изменение потенциала происходит по всей поверхности клетки, что отражает собой уменьшение или усиление заряда в клетке. Специальными исследованиями Элула и сотрудников, было показано, что на поверхности каждой нервной клетки имеется несколько участков, являющихся источниками волновой активности. Так, например, если измерить уровень генерации спайков для спонтанно разряжающегося нейрона (рис. 5), то хорошо заметны вариации этого уровня (пунктирная линия) в несколько милливольт. В то же время известно, что при прямом электрическом раздражении коркового нейрона порог возникновения спайка, как правило, постоянен.
Необходимо также подчеркнуть, что Элул, рассматривая роль глиальных клеток как возможного источника внеклеточного тока и их вклада в образование поверхностной ЭЭГ, полагает, что этот вклад не является решающим, по сравнению с активностью нервны:: клеток, Отрицал роль глин в электрогенезе ЭЭГ также М. Н. Жадин (1973).
Поэтому, как полагает Элул, такую вариацию порога v спонтанно разряжающегося нейрона можно объяснить различным сопротивлением между кончиком микроэлектрода и разными источниками последовательных волн, возникающими в разных участках мембраны нейрона. Исходя из того, что сопротивление пропорционально расстоянию между микроэлектродом и синапсом, одинаковый ток от двух синапсов, расположенных на разных участках мембраны, может регистрироваться как неодинаковый. Таким образом, поверхность клетки не является эквипотенциальной, что не соответствует предположению образования волн только за счет понижения или повышения общего заряда клетки.
Элул приходит к выводу, что унитарными генераторами волновой активности являются отдельные синапсы, а волны, наблюдаемые при внутриклеточных регистрациях, представляют собой уже суммарный выход этих синаптических вкладов.
Однако, исходя из того, что пресинаптические волокна ветвятся и образуют множество синаптических контактов с каждым постсинаптическим нейроном и что эти синапсы активируются почти одновременно при срабатывании пресинаптического волокна, автор указывает, что «унитарный генератор скорее всего должен быть представлен группой синапсов, имеющих общий пресинаптический вход, чем отдельным синапсом. Поэтому можно ввести понятие синаптической функциональной единицы по аналогии «с моторной» единицей в скелетной мышце.
Таким образом, «синаптические функциональные единицы» можно рассматривать как элементарные, унитарные генераторы волновой активности. Именно они должны быть той единицей активности, из которых «строится» суммарная макро-ЭЭГ. Однако вопрос о том, какая часть нейрона (сома, базальные или апикальные дендриты) играет основную роль в образовании суммарной ЭЭГ того или иного отдела мозга, остается еще не совсем ясным.
Надо, видимо, полагать, что вклад дендритных образований в подавляющем большинстве случаев более весом, чем сомы нейронов, хотя, как указывает Элул, в связи со сложной топологией (геометрией) внеклеточного пространства коры больших полушарий, затрудняющего интерпретацию внеклеточных измерений активности, трудно установить, апикальные или базальные дендриты являются источником волновой активности.
В некоторой степени это упрощается при работе на других структурах мозга (таких, например, как гиппокамп), где точно можно сказать, откуда производится регистрация — из сомы нейрона, апикальных или базальных дендритов.
Кроме того, признавая преимущественно синаптическое происхождение волновой нейрональной активности, нельзя, видимо, сбрасывать со счета «нестабильность» возбудимых мембран, т. е. способности их при определенных условиях («следствие деполяризации, изменения концентрации кальция и т. д.) генерировать осцилляции. В «том случае, как указывает Элул, синапсы либо вызывают, либо подавляют эти осцилляции, играя только ключевую роль регуляторов, хотя и не являются первичным источником волновой активности. К такому «осциллирующему» поведению, видимо, более способны тонкие дендритные ветви.
➥ Основная статья: Физиология дендритов нервных клеток и их функциональная роль
Связь между ЭЭГ (макроактивностью) и унитарными генераторами
Ритмические токи, генерируемые отдельными нейронами и идущие через мембраны во внеклеточную среду, могут взаимно складываться или уничтожаться.
Элул (Elul, 1972) рассмотрел биофизические факторы, имеющие отношение к суммированию отдельных генераторов. Он полагает, что унитарные генераторы, расположенные рядом с отводящим электродом, должны иметь больший удельный вес в регистрируемом потенциале, чем генераторы, расположенные дальше; причем их электрические поля должны суммироваться линейно. Иными словами, удвоение числа активных генераторов в данном объеме ткани должно приводить и к удвоению величины активности, регистрируемой макроэлектродом с поверхности коры (т. е. на определенном удалении от активных генераторов).
Такое линейное суммирование возможно лишь при допущении, что внеклеточная среда однородна. Однако, как показывают данные Элула (Elul, 1962), Калвета и сотрудников (Calvet et al., 1964), распространение активности в коре не является однородным во всех направлениях. Такая неоднородность внеклеточной среды определяется геометрией внеклеточного пространства и его емкостными свойствами, что, видимо, в определенной степени затрудняет оценку участия количества генераторов в образовании ЭЭГ в той или иной точке отведения. И, наконец, нелинейность привносится регистрирующим суммарную ЭЭГ электродом, который имеет неоднородную чувствительность к дипольным генераторам с различным их пространственным расположением (Peronnet et al., 1971).
Так, например, было показано, что в зависимости от местоположения диполей, даже четко ориентированных по отношению к плоскости электрода, электрод ряд диполей вообще «не замечает», а ряд диполей более, удаленных вносит больший вклад в потенциал у электрода, чем, например, диполи, находящиеся ближе, но менее благоприятно расположенные.
Таким образом, даже для такой структуры, как кора больших полушарий, где имеет место четкая ориентация большого количества клеточных элементов (а значит, и ориентированных диполей) трудно дать четкое определение популяции нейронов, дающих свой вклад в тот или иной суммарный потенциал.
Если же представить себе более случайную ориентацию диполей (например, ядерные мозговые структуры, не имеющие четкой ориентации клеточных элементов), то для оценки вклада нейронной популяции в суммарный потенциал требуется, видимо, статистический подход. Как правило, система с хаотично ориентированными диполями обычно и не приводит к возникновению высокоамплитудной ЭЭГ. В связи со всеми этими ограничениями Элул (1966) полагает наиболее правильным рассматривать вопрос суммирования активностей генераторов статистически.
Статистический подход к анализу связи между спонтанной макро-ЭЭГ и волновой активностью отдельных нервных клеток показал (Elul, 1972), что в основе спонтанной макроактивности лежит суммирование синхронизированной активности относительно небольшой части нейронной популяции мозговой ткани. Остальная часть популяции нейронов также активна, но отдельные элементы ее работают по синхронно и поэтому суммируются менее активно.
Чтобы более четко в этом разобраться, необходимо хотя бы кратко коснуться основных доказательств, приводимых Элулом.
Теоретически можно рассмотреть две крайние ситуации. Если популяция генераторов, участвующих в образовании активности, регистрируемой на поверхности, будет полностью синхронизированной, то макро-ЭЭГ синхронна с волновой активностью любого генератора. Когда же отдельные генераторы работают независимо друг от друга, то в этом случае связь между элементарными генераторами носит случайный характер и любой одиночный генератор не будет синхронным с ЭЭГ. Казалось бы, настолько разные механизмы должны дать и различный конечный результат. Однако интересно то, что результат в обоих случаях будет очень сходным.
Теоретически Элул показал, что если связь между отдельными генераторами носит случайный характер, то в этом случае их суммарная активность приводит к генерации суммарной активности, имеющей такой же общий вид и частотное содержание, что и активность любого отдельного генератора. Однако синхронизации суммарной активности с. любым отдельным генератором не наблюдается.
Видимо, экспериментально этот вопрос можно решить лишь в опытах с внутриклеточной регистрацией активности корковых нейронов. При внутриклеточной регистрации каждая «синаптическая функциональная единица» (унитарный генератор) может работать независимо от других таких единиц нейрона. В этом случае каждый нейрон может представлять собой активность многих тысяч унитарных генераторов (учитывая, что число синапсов на нейроне может составить 10³, Косицын, 1964). Тогда в результате случайной связи между такими генераторами внутриклеточная волновая активность будет смесью отдельных синаптических потенциалов и лишь случайно связана с активностью того или иного генератора и не будет синхронной с суммарной ЭЭГ. Если генераторы работают синхронно, то внутриклеточная запись должна быть синхронна с суммарной ЭЭГ.

Отмечали, что действительно существует положительная корреляция между внутриклеточной активностью и спонтанной ЭЭГ. Однако Элул такой устойчивой синхронности и корреляции не обнаружил. Анализируя эти противоречия, он нашел, что положительная корреляция, хотя и имеет место, очень непродолжительна—до 1 с (Elul, 1966, 1968, 1972). Так, на рис. 6 видно, что на коротком отрезке времени (ограниченном вертикальными штриховыми линиями) внутриклеточная активность почти совершенно синхронна с ЭЭГ. Однако за этими границами такой связи не наблюдается. Статистически показано отсутствие для долгих периодов активности, устойчивой корреляции между ЭЭГ и внутриклеточной активностью. Специальные исследования, предпринятые Элулом, показали, что стойкой синхронизации, фиксированной группы генераторов с суммарной ЭЭГ, не происходит, т. е. «в коре нет специфического класса специализированных генераторов ЭЭГ».
Полученные данные позволили сделать вывод, что «все нейронные генераторы попеременно играют эту роль временами синхронизируясь с ЭЭГ. Синхронизация захватывает то одну, то другую группу нейронов в течение долей секунды». Иными словами, спонтанная ЭЭГ, регистрируемая с поверхности коры, представляет собой прерывистую синхронизацию сравнительно небольших групп нейронов. Автор предположил, что прерывистая синхронизация нейронных групп в последовательные моменты времени осуществляется в коре благодаря специальным влияниям из подкорковых структур, где имеет место пейсмекер (водитель) такого ритма.
Каковы же причины «спонтанности» суммарной активности мозга?
Как мы уже говорили, впервые обратил внимание на спонтанные колебания биопотенциалов И. М. Сеченов. В последующие годы термин «спонтанные колебания» широко используется в. электрофизиологии. Спонтанную активность ЦНС (особенно высших отделов мозга) и ее кажущуюся независимость от афферентных влияний ряд авторов (Brown, 1914; Bullock, 1957, и др.) рассматривали как принципиальную основу функционирования нервной ткани.
Независимость электрической спонтанной активности от внешних влияний была найдена в опытах многочисленных исследователей. Так, спонтанная суммарная активность в изолированном участке мозга была впервые описана у амфибий (Gerard, Joung, 1937). Эти колебания могли наблюдаться в течение нескольких часов, что привело авторов к предположению, что и нейроны целого мозга могут проявлять спонтанную активность при отсутствии афферентных нервных импульсов. Бремер, Фессар и другие исследователи также полагали, что источник спонтанной активности находится в самих нейронах. Эдриан и Бьютендайк (Adrian, Buylendijk, 1931) на препарате головного мозга рыбы нашли четкую спонтанную активность в продолговатом мозге.
У различных классов и видов беспозвоночных в ЦНС также была найдена спонтанная электрическая активность, которая возникала и в изолированных участках нервной ткани последних в течение многих часов (Adrian, 1931; Rijlant, 1931; Prosser, 1946, и многие другие).
Естественно, возник вопрос об аутогенной, истинно спонтанной, природе этих колебаний. Однако еще И. М, Сеченов, а затем ряд последующих исследователей (Burns, 1958; Смолин, 1962, и др.) справедливо указывали, что биоэлектрические колебания мозга не являются в истинном смысле спонтанными, — они детерминированы трудно учитываемыми факторами, такими, как самые различные интеро- и экстероцептивные воздействия, колебания температуры, интенсивность обмена веществ и т. д.
С точки зрения многих исследователей спонтанная активность возникает исключительно под влиянием синаптической активации. Опыты с изолированными участками нервной системы, о которых шла речь выше, объяснялись наличием постоянного источника раздражения с места повреждения нервной ткани (края разреза). Поэтому возникла тенденция замены термина «спонтанные колебания»— термином «фоновые колебания» биопотенциалов. В то же время такая замена термина, видимо, не решала вопроса относительно сути явления спонтанности. Необходимо было более четко определить термин «спонтанность». В ее зависит от того, какой смысл исследователь вкладывает в этот термин. В последнее время для подавляющего большинства исследователей этот термин отнюдь не подразумевает признание индетерминизма тех или иных процессов в организме.
Говоря о «спонтанности» тех или иных процессов (применительно к биоэлектрическим процессам мозга) подчеркивается лишь тот факт, что эти процессы находятся «в ином причинно-следственном ряду событий по сравнению с процессами, более непосредственно связанными с текущими воздействиями на организм».2
Иными словами, не отрицая факта четкой детерминированности того или иного «спонтанного» процесса и даже вскрывая причины и тонкие механизмы такой «спонтанности», мы не всегда находим связь этих процессов с текущей нервной деятельностью. И действительно, имеется достаточно факторов как в рецепторах (Гранит, 1957), так и в периферической и ЦНС, которые могут привести к спонтанной активности нейронов (и, естественно, суммарной активности), не связанной с текущей нервной деятельностью и информационными процессами (хотя и влияющей на них). Такая спонтанная активность нейронов может быть связана с колебаниями температуры, интенсивностью обмена веществ, со случайными выделениями квантов медиатора в синапсах, достаточных для достижения критического уровня спайка и с другими факторами. Экспериментально существование истинно спонтанных ритмов доказано для мышечных клеток (Шаповалов, 1966), но это труднее сделать в отношении нервных клеток, так как нельзя без повреждения изолировать их или избирательно блокировать только их хемо- рецептивную мембрану. Однако Хилд и Тасаки (Hild, Tasaki, 1963) обнаружили спонтанную ритмическую активность в нервных клетках культуры ткани мозжечка кошек и крыс, в клетках, лишенных синапсов. Это, видимо, безусловно доказывает возможность аутогенной активности у нервных клеток. Поэтому можно полагать, что фоновая, в частности импульсная, активность мозга состоит из двух компонентов. Один из них должен отражать какие-то циклические координированные процессы в нервной сети, а другой — истинную форму спонтанной импульсной активности, прямо не связанную с этими процессами.
Известно (Костюк, 1965), что импульсная активность в любом нейроне возникает вследствие деполяризации его мембраны под влиянием как синаптического возбуждающего действия, так и многих несинаптических факторов. В первом случае ритм генерации спайка нейроном задается ритмом градуальной деполяризации. Иными словами, к нейрону поступает периодически возбуждающая импульсация, и деполяризация его мембраны в этом случае также носит волнообразный характер и каждый раз при достижении критического уровня возникает слайдовый разряд. Во втором случае такие трансмембранные колебания разности потенциалов почти незаметны; разность потенциалов на мембране происходит постепенно (например, при медленном течении метаболических процессов в клетке), однако клетка периодически генерирует импульсы с определенной частотой соответственно с уменьшением трансмембранной разности потенциалов. В этом случае ритм нейрона является аутогенным. Такая импульсная активность нейрона может быть довольно стойкой, без каких-либо признаков аккомодации. Она наблюдается в различных промежуточных и двигательных нейронах спинного и головного мозга.
А. И. Шаповалов (1966) также указывает, что данные, полученные при различных способах активации клеток, позволяют провести параллель между фоновой активностью и аутогенной ритмикой. Во всех случаях постоянная постсинаптическая деполяризация вызывает в нейроне локальные процессы деполяризации, которые при достижении критического уровня генерируют потенциалы действия. Однако при определенных условиях локальные процессы деполяризации, ведущие к спонтанной ритмике, видимо, могут возникать и при отсутствии специальных пресипаптических воздействий, т. е. приводить к истинной аутогенной ритмике (например, спонтанные разряды мотонейрона в среде с пониженным количеством ионов кальция).
К элементам, обнаруживающим спонтанную активность, относятся нейроны ряда зон коры и особенно двигательной зоны, таламуса, гиппокампа, ретикулярной формации ствола и нейроны ряда других нервных центров.
Внутриклеточные отведения от нейронов новой коры свидетельствуют, что их спонтанная активность характеризуется колебаниями мембранного потенциала, причем, как мы отмечали ранее, спонтанные ВПСП, часто достигающие огромной величины, могут не сопровождаться спайковыми разрядами или, в более редких случаях, спонтанная активность корковых нейронов может выражаться в спонтанных ТПСП. И, наконец, спонтанная активность может быть выражена в виде спайков, которым, как правило, предшествует медленная волна деполяризации.
Однако до сих пор еще не решен вопрос, свойственна ли нейронам коры больших полушарий, хотя бы в какой-то мере, спонтанная, аутогенная ритмика. С этой целью были предприняты многочисленные попытки в работе на нейронах изолированных полосок коры и изолированной коре, где спонтанная активность нейронов хорошо выражена. Естественно, что решить этот вопрос для нейронов сложных нервных сетей п особенно таких, как новая кора, крайне трудно, так как всегда сохраняется возможность того, что фоновая активность данного нейрона определяется импульсацией с соседних нервных образований.
Таким образом, фоновая активность нейронов может быть аутогенной и вызванной, механизмы их, видимо, столь многогранны, что требуют рассмотрения в каждом отдельном случае.
Аутогенная и синаптически вызванная активность нейрона (Шаповалов, 1962, 1966) могут, вероятно, находиться в сложных отношениях друг с другом и играть важную роль в его интегративных функциях. Возможно также, что при изучении закономерностей течения метаболических процессов в организме, гормональной регуляции или других процессов такая «аутогенность» нейронов в ряде случаев уложится в определенные рамки четких рефлекторных причинно-следственных отношении, где термин «спонтанность» будет уже не уместен.
Какие же общие механизмы лежат в основе синаптически вызванной фоновой нейронной активности (и суммарной активности) в условиях полного покоя животного или человека и в отсутствие внешних раздражении?
Прежде всего, она обусловлена постоянной синаптической бомбардировкой нейронов притоком афферентных импульсов от различных рецепторов и других нейронов. Этот приток афферентации из сенсорных систем является существенным фактором, влияющим на формирование фоновой активности.
В многочисленных исследованиях на животных было четко показано, что лишение нервного центра притока афферентной сигнализации приводит к частичному или полному исчезновению фоновой биоэлектрической активности, либо к резкому изменению ее частоты. Таких данных можно привести большое количество. Все они свидетельствуют, что рецепторные образования, особенно зрительный и вестибулярный аппараты, могут рассматриваться как стимуляторы, поддерживающие тонус ЦНС. Сами рецепторные образования обладают «спонтанной» активностью (Гранит, 1957), являющейся основным источником активация вышележащих нервных центров.
При физиологическом покое и при отсутствии каких-либо раздражений, кроме синаптической бомбардировки от рецепторов, имеют место и постоянные влияния на мозг афферентных импульсов от рецепторов внутренних органов, сосудистой системы, и, возможно, от ряда нервных клеток, обладающих аутогенной спонтанной активностью и имеющих место в дыхательном центре, перегородке и, возможно, в ряде других нервных центров. Вся эта импульсация играет определенную роль в провокации и поддержании фоновой активности нейронов и тем самым суммарной фоновой активности. Известна, например, активирующая роль спонтанной активности дыхательного центра, оказывающей определенное влияние на центры спинного и головного мозга. Кроме того, не исключено, что спонтанная активность мозга может быть обусловлена длительной циркуляцией возбуждения (Lorente de No, 1938) по замкнутым цепям нейронов.
Таким образом, механизмы фоновой активности мозга, видимо, крайне многообразны и до конца еще не исследованы. Тем не менее возникает принципиальный вопрос— каково функциональное значение фоновой активности головного мозга, какую роль она может играть в интегративных функциях ЦНС? Мнения исследователей здесь расходятся: одни рассматривают фоновую активность как шум в нервной системе (например, Wall, 1959), другие полагают, что фоновая электрическая активность мозга играет важную роль в интегративных функциях на разных уровнях ЦНС.
Так, на уровне отдельных нейронов спинного мозга изучение взаимодействия спонтанной и синаптической вызванной активности привело А. И. Шаповалова (1966) к выводу, «что такое взаимодействие имеет определенное значение во взаимной координации деятельности различных нейронов и играет роль высокочувствительных фильтров в нейронных сетях».
Функциональное значение фоновой активности нейронов заключается, по мнению ряда авторов (Гранит, 1967; Jung, 1961, и др.), в обеспечении возможности ответа системы и отдельных нейронов как в сторону торможения активности, так и в сторону ее усиления. Нейроны, лишенные фоновой активности, активируются только в одном направлении. Можно полагать, что нейрон, обладающий фоновой активностью, является более чувствительным передатчиком информации, чем молчащий нейрон. Наличие фоновой активности способствует высокому уровню дифференциальной чувствительности и гибкости системы и позволяет избежать фиксированных порогов.
Ряд исследователей считает, что фоновая активность способствует высокой чувствительности рецепторов к внешним воздействиям. Определенное значение придается тонической активации со стороны спонтанно активных нейронов для поддержания определенного тонуса нервных центров, что прямо связано с механизмами переработки информации.
Таким образом, если существуют экспериментальные данные, что фоновая активность нейронов тесно связана с их интегративной деятельностью, то несколько сложнее обстоит дело, когда это касается спонтанной суммарной фоновой активности мозга и ее функционального значения. С этой точки зрения наибольший интерес представляют спонтанные ритмические электрические суммарные колебания, где apriori можно сказать, что они должны отражать довольно синхронное возникновение в огромных популяциях нейронов коры возбудительных или тормозных колебаний потенциала. Такая синхронизация может охватывать огромное или подавляющее большинство нейронов не только одной нервной структуры, но одновременно вовлекать и ряд других нервных центров.
Способность головного мозга человека и животных генерировать регулярные, ритмичные суммарные электрические колебания неоднократно привлекала к себе внимание многих исследователей, поскольку с полным основанием можно было надеяться, что вскрытие внутренней природы этого интересного явления даст много нового в понимании механизмов деятельности мозга.
Зависимость параметров двигательной реакции человека от фазы электрической активности мозга, на которую приходится стимул, зависимость различения параметров сигнализации от характера ритмической активности мозга, указания на возможную связь суммарных ритмических колебаний с отсчетом времени, с ориентировочной и условнорефлекторной деятельностью, с механизмами памяти и эмоций, а также возникновение генерализованной ритмической активности при естественном и наркотическом сне и некоторых патологических состояниях и т. д. показывают, что изучение механизмов генерации спонтанной ритмической электрической активности может внести существенный вклад в понимание ее функционального значения.
Рассмотрим некоторые общие точки зрения на происхождение спонтанной ритмической электрической активности головного мозга.
Гипотезы происхождения спонтанной ритмической электрической активности головного мозга
В гипотезах о происхождении ритмической активности мозга ряд авторов предполагает, что электрическая активность мозга вообще, а следовательно, и ритмические виды активности, являются отражением процессов, не имеющих прямого отношения к сигнальной, информационной деятельности мозга (Gibbs, 1938; Артемьев, 1962, и др.). Однако последующие исследования показали, что более правы были те исследователи, которые связывали возникновение ритмической электрической активности непосредственно с определенной картиной состояний возбуждения и торможения в нейронах. Одной из первых в этом направлении можно считать гипотезу Эдриана и Мэтьюза. Изучая спонтанную ритмическую электрическую активность мозга человека (α-ритм) и изменение этой активности под влиянием ритмических световых раздражений, они выдвинули предположение, что частота разрядов корковых нейронов синхронно периодически изменяется. Когда частота разрядов и количество разряжающихся нейронов увеличивается, суммация их потенциалов действия (импульсных разрядов) приводит к появлению на поверхности коры медленной электрической волны; при спаде частоты нейронных разрядов наблюдается спад волны и такие изменения происходят периодически с частотой α-ритма. В настоящее время это предположение имеет лишь исторический интерес, прежде всего потому, что оно не объясняет, за счет чего может происходить синхронизация нейронной активности, не говоря уже о том, что предположение об образовании медленной электрической активности мозга путем суммации импульсных потенциалов действия не было подтверждено. Однако гипотеза Эдриана и Мэтьюза выдвинула важный вопрос о природе нейронной синхронизации, т. е. об одновременной деятельности большого числа нейронов, которая, очевидно, необходима для генерации значительных суммарных токов и колебаний потенциалов, регистрируемых в виде ритмической электрической активности.
Гипотеза Лоренте де Но (Lorente de No, 1938) —это первая попытка объяснить способность нейронов к синхронной деятельности на основе особенностей их взаимных связей. Исследования Лоренте де Но показали, что взаимосвязи между нейронами могут образовывать замкнутые кольца. Поэтому предполагалось, что возникшее в такой замкнутой цепочке возбуждение будет передаваться от нейрона к нейрону и сможет циркулировать в ней значительное время. При этом любой нейрон цепочки будет давать периодические разряды, интервал между которыми определяется временем прохождения возбуждения по цепи.
Эта гипотеза не получила подтверждения в дальнейших физиологических исследованиях. Сейчас ясно, что она имеет целый ряд пробелов (трудно объяснить возникновение синхронных разрядов значительного количества нейронов, сохранность компактности залпа импульсов, циркулирующего по длинной цепи, и как циркуляция такого залпа находит отражение в медленных электрических колебаниях и т. д.). Высказывания о возможности длительной передачи импульсов по нейронным цепочкам встречаются и у других исследователей (Binns, 1951; Verzeano et al., 1954, 1960, и др.), но они уже не приписывают им основную роль в генерации медленной ритмической активности.
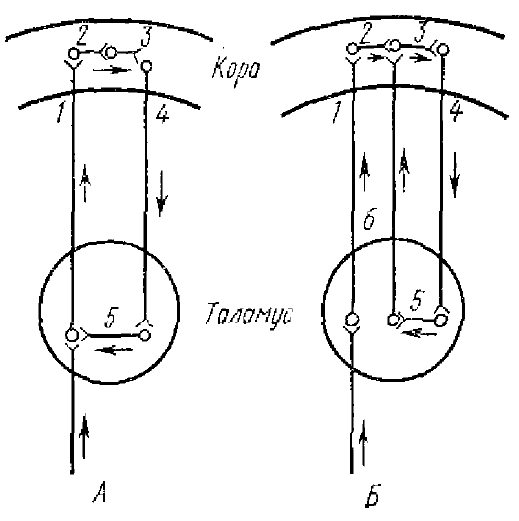
1, 2, 3, 4, 5, 6 — схематическое изображение нейронов, составляющих кортико-таламические круги. Стрелки — направление циркуляции импульсов
Вариантом гипотезы Лоренте де Но явилась гипотеза таламо-кортикальных кругов, предложенная Чангом (Chang, 1950), которая основана на данных, показывающих, что кроме основных таламо-кортикальных связей можно обнаружить также и связи обратного направления— кортико-таламические. Чанг показал, что разряды последействия синхронно возникают в различных проекционных зонах коры и соответствующих им проекционных ядрах таламуса как при афферентном, так и при электрическом раздражении коры. В отличие от других исследователей он утверждал, что для генерации разряда последействия равно необходимы как кора, так п таламические ядра. Разряд последействия не может быть вызван ни в коре после разрушения проекционного таламического ядра, ни в таламусе после разрушения коры или перерезки радиации. Отсюда он сделал вывод, что основную роль в генерации ритмической активности играет взаимодействие таламо-кортикальных и кортико-таламических связей: их комбинация образует замкнутое кольцо, в котором может происходить циркуляция возбуждения, выражающаяся в ритмических колебаниях потенциала, регистрируемых от коры и таламуса (рис. 7).
Однако основные факты, на которых строится изложенная гипотеза, ставятся под сомнение. Данные об исчезновении ритмического разряда последействия в таламусе после удаления коры или пересечения таламокортикальных связей не только противоречат более ранним данным (Adrian, 1941; Morison, Dempsey, 1942), но и не подтвердились последующими работами (Bremer, Bonnet, 1950; Adrian, 1951). Поэтому большинство авторов отвергли гипотезу Чанга.
Гипотеза Клэра и Бишопа (Clare, Bishop, 1956). Авторы полагали, что в основе образования ритмической активности играют основную роль свойства каждого отдельного нейрона, точнее, его синапсов. Исследуя циклы восстановления различных вызванных потенциалов (ВП) коры, Клэр и Бишоп обнаружили, что кривые восстановления для некоторых видов вызванных потенциалов (включая потенциалы классической реакции вовлечения, см. лекцию 6) имеют сложный характер, В начале (около 15 мс после обусловливающего стимула) наблюдается экзальтация тестирующего ответа, затем — более или менее глубокая депрессия, длящаяся 1/6—1/8 с. По окончании периода депрессии наблюдается фаза повторной супернормальности, после которой тестирующий ответ возвращается к исходному уровню. Основное значение для возможности установления ритмической активности нейронов имеют фазы депрессии и повторной супернормальности. Эта повторная экзальтация может быть настолько сильной, что порог возникновения вызванного потенциала в это время падает практически до нуля, т. с. следующий потенциал мог возникать и без раздражения, самопроизвольно. Таким путем, очевидно, может возникнуть и спонтанная ритмическая активность. При ритмических раздражениях максимальный эффект будет наблюдаться в том случае, если каждый стимул приходится на фазу вторичной супернормальности от предыдущего стимула, т. е. при частоте раздражения 6-8 в 1 с, что и наблюдается действительно в виде реакции вовлечения. Повторная активация нейрона раньше, чем через 1/6—1/8 с после предшествующей, маловероятна, так как в эго время он еще находится в фазе депрессии. Поэтому все нейроны будут разряжаться периодически с частотой 6—8 залпов в 1 с, что, естественно, должно создать благоприятные условия для объединения их активности в общем ритме (например, за счет взаимной синаптической бомбардировки).
Описанная характерная форма циклов возбудимости нейронов, определяемая по показателю вызванных потенциалов, является общераспространенным явлением. Данные, согласующиеся с результатами Бишопа и Клэра, были получены рядом авторов для различных видов вызванных потенциалов. В большинстве случаев длительность фазы депрессии до начала вторичной супернормальности составляет около 100 мс, т. е. близка к периоду ритмической активности коры и таламуса — α-ритма и веретенообразной активности.
Что касается природы циклов возбудимости описанного типа, то, по предположению Бишопа и Клэра, они являются отражением свойств одного из видов синапсов, располагающихся на дендритах корковых нейронов. Предполагается, что возбудимость этих синапсов после однократной активации меняется во времени по типу экзальтация — депрессия — экзальтация, что и обнаруживается по вызванным потенциалам коры.
Бишоп и Клэр основную роль в формировании реакции вовлечения и аналогичной ей спонтанной активности приписывают синапсам корковых нейронов. Это противоречит фактам, которые показывают, что первичным очагом этих видов ритмической активности являются таламические ядра. Основной аргумент — появление реакции вовлечения не только при раздражении таламуса, но и при раздражении коры или радиации — не вполне состоятелен, так как при этом возможна антидромная активация таламических нейронов, в которых и может происходить процесс рекрутирования, отражающийся затем вторично в потенциалах коры мозга. Однако это не является принципиальным в гипотезе Бишопа и Клэра. Она вполне может быть применена и к таламическим нейронам, для которых также описаны циклы возбудимости, включающие довольно длительный период депрессии и период вторичной экзальтации (Bishop, O’Leary, 1940, Andersen, 1964).
Основной недостаток гипотезы Бишопа и Клэра заключается в том, что свойство нейронных синапсов менять свою проводимость после однократного возбуждения только постулировалось, но не имело прямых экспериментальных доказательств. В то же время эта гипотеза уже имела под собой существенную экспериментальную базу, поскольку, с одной стороны, описанный выше тип восстановления реактивности нервных элементов (каков бы ни был его механизм), безусловно, существует, а с другой — он, очевидно, должен действительно способствовать ритмической синхронизации работы нейронов именно таким образом, как предполагалось в разбираемой гипотезе.
Дальнейшая разработка теории происхождения ритмической активности мозга связана с исследованием синаптической организации структур, генерирующих” такую активность. Исследованиями многих авторов установлены подробные картины синаптических процессов в различных таламических ядрах, наблюдаемые как во время спонтанной ритмической активности, так при однократном их возбуждении, и при ритмических раздражениях. Существенно, что как при одиночном, так и при ритмическом возбуждении таламических ядер картины синаптических процессов, наблюдаемые после каждого стимула, количественно могут существенно различаться, по качественно сходны по ряду признаков.
Прежде всего, в ответ как на прямое, так и на афферентное раздражение таламического ядра наблюдается относительно короткий (15—20 мс) ВПСП, сопровождающийся спайковыми разрядами нейронов (Purpura et al., 1963, 1965; Andersen et al,, 1964, и др.) (см. рис. 8). Вслед за ВПСП в таламических нейронах возникает ТПСП большой длительности (порядка 100 мс и более), обнаруживающийся как по экстраклеточным потенциалам, так и при. внутриклеточной регистрации, а также, по угнетению, в это время импульсной активности клеток. Исследование природы этого ТПСП показало, что он обусловлен деятельностью возвратных тормозных связей через коллатерали аксонов таламических нейронов (см. рис. 9); такой же ТПСП возникает и при антидромной активации таламического ядра (Vastola, 1959; Andersen et al., 1964). ТПСП в нейронах наблюдались и без предшествующих спайковых разрядов; это показывает, что торможение таламических нейронов может быть вызвано и деятельностью соседних с ними нейронов, т. е. является возвратным торможением.
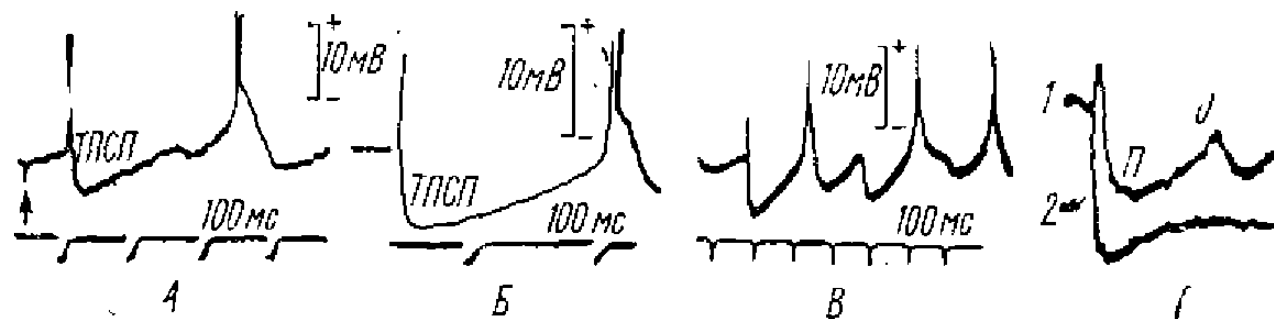
П — положительная волна и О — отрицательная волна
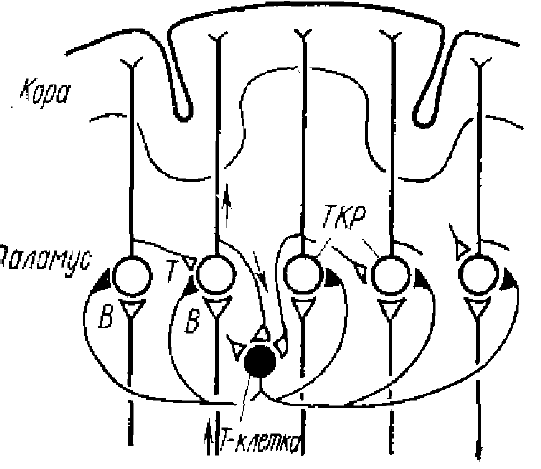
Существование в таламических нейронах возвратного коллатерального торможения не. является свойством только этих нейронов. Оно описано почти для всех уровней анализаторов: на рецепторном уровне, в сетчатке, в клетках клиновидного ядра, а выше таламического уровня — для корковых нейронов. Такое торможение описано также в старой коре мозга, в коре мозжечка, в спинном мозге для мотонейронов. Нами оно найдено у низших позвоночных в обонятельной луковице и коре черепах. Не исключено, что в центральных звеньях анализаторов это торможение играет важную роль. Доводом в пользу такого предположения может служить прямое его участие в установлении ритмической синхронизации нейронной активности.
Большая длительность возвратных ТПСП таламических и корковых нейронов наводит на мысль, что они могут иметь отношение к формированию ритмической активности мозга.
Как показали исследования (Purpura, Cohen, 1962; Purpura, Shofer, 1963; Andersen et ah, 1962, 1964), длительные ТПСП являются в таламических ядрах обязательным компонентом как спонтанной веретенообразной активности, так и вызванных электрическим раздражением разрядов последействия или реакции вовлечения. При этом внутриклеточно регистрируемые ТПСП совпадают с медленными позитивными электрическими колебаниями, регистрируемыми при макроэлектродном экстраклеточном отведении, так что возникновение этих колебаний в результате суммации синхронно возникающих в таламических нейронах ТПСП едва ли может вызвать сомнение.
Совпадение длительности ТПСП таламических нейронов с периодом спонтанной ритмической веретенообразной активности может навести на мысль о доминирующей роли ТПСП в установлении ритмической активности нейронов. Этот шаг был сделан Андерсеном, Экклсом и соавторами, которые в серии работ экспериментально разработали и обосновали теорию происхождения ритмической синхронизации нейронов.
Footnotes
- Соколов Е. Н. Природа фоновой ритмики коры больших полушарий. — В сб.: Основные вопросы электрофизиологии центральной нервной системы. Киев, 1962, с. 178.
- Гращенков Н. И., Латаш Л. П., Файгенберг И. М. Диалектический материализм и некоторые проблемы современной нейрофизиологии-—В сб.: Философскис вопросы физиологии высшей нервной деятельности. М., Изд-во АН СССР, 1963. с. 54.