Дендриты нервной клетки до сих пор являются наименее изученными. Это прежде всего связано с тем, что невозможно ввести микроэлектрод внутрь дендрита большинства нервных клеток, а косвенные методы изучения его функциональных особенностей не всегда могут дать необходимый результат.
Поэтому исследователи обращаются к суммарным электрическим реакциям мозга, происхождение которых они связывают с активностью дендритов. Так, при изучении суммарных электрических ответов коры больших полушарий на раздражение периферических структур и особенно при изучении ответа коры на ее прямое раздражение ряд авторов (Chang, 1951; Ройтбак, 1955, 1968; Ochs, 1962, и др.) пришли к выводу, что эти ответы коры частично или целиком обусловлены активностью апикальных дендритов пирамидных клеток. Ответ коры на ее прямое раздражение получил название «дендритный потенциал». На основании его изучения многие исследователи пришли к выводу, что дендриты обладают мембраной, которая по своим свойствам резко отличается от свойств мембран тела клетки и аксона, и высказаны разные точки зрения о способности дендритов корковых нейронов к проведению возбуждения по типу «все или ничего».
Так, в работах Пурпуры (1963), Окуджавы (1963), Грундфеста (1964) и др. был сделан вывод, что дендритные потенциалы являются локальными, градуальными потенциалами; сама же дендритная мембрана неспособна к активному проведению возбуждения. Однако Чанг (Chang, 1952) считал, что раздражение поверхности коры вызывает возбуждение дендритов, которое распространяется со скоростью 1—2 м/с. Бишоп и сотрудники (Clare, Bishop et al., 1955; Bishop, 1958) сделали вывод, что распространение возбуждения по дендритам возможно только в антидромном направлении. Показателем такого антидромного проведения, т. е. проведения возбуждения по дендритам от клеточных тел к периферии дендрита, они считали двухфазный положительно-отрицательный потенциал, регистрируемый с поверхности коры при раздражении ее глубоких слоев. При ортодромном раздражении. т. е. под влиянием синаптически вызванной деполяризации, в дендритах, по их мнению, возникают местные «пиковые» потенциалы, распространяющиеся с затуханием и принимающие участие в дендро-соматическом проведении возбуждения.
Вопрос о возможности активного проведения возбуждения по дендритам нервных клеток является принципиальным с точки зрения их роли в обработке информации, поступающей к нейрону: нейроны с пассивным проведением возбуждения по дендриту представляют собой лишь нейроны-сумматоры; при активном проведении по дендритам им отводится роль нейронов, анализирующих приходящую афферентацию на основе выполнения элементарных логических функций (Богданов, Головчинский,1970).
К настоящему времени в литературе накопились прямые и косвенные экспериментальные факты, показывающие, что нет, видимо, принципиальной разницы между свойствами мембраны дендритов и остальной мембраной нейрона. Во многих работах показано, что мембрана дендритов (по крайней мере, мембрана толстых дендритных ветвей) способна проводить импульсы по типу «все или ничего», т. е. спайковую активность. Например, у спинального мотонейрона базальные наиболее утолщенные части дендритов могут генерировать пиковый потенциал. Полагают, что импульс проводится в разветвления дендрита. Так, Фэтт (Fatt, 1957). изучая распределение электрических полей в момент антидромного разряжения одиночного мотонейрона кошки, пришел к выводу, что возбуждение распространяется по дендритам до 300 мкм от тела клетки со скоростью 0,7—1 м/с.
Терцуоло и Араки (Terzuolo, Araki, 1961) при изучении активности мотонейрона кошки двумя микроэлектродами показали, что антидромное возбуждение приводит к появлению потенциала действия сначала в соме, а затем в дендрите. Этот факт также показал, что возбуждение с сомы клетки распространяется, как пишут авторы, «хотя бы по некоторой части дендритов». Близкие данные получил Рамос (Ramos, 1969). Распространение потенциала действия по дендритам убедительно доказано Хильдом и Тасаки (Hild, Tasaki, 1962), работающим на культуре нервной ткани мозжечка. Они имели возможность раздражать и регистрировать активность дендритных ветвей крупных нейронов ткани, таких, как клетки Пуркинье, на расстоянии до 100 мкм от сомы клетки. Показано. что возбуждение может распространяться по дендритам в обоих направлениях со скоростью 0,1 м/с.
При внутриклеточной регистрации на животном и естественной синаптической активации нейронов выяснить способность дендритов к генерации и проведению потенциала действия крайне сложно. В этом случае необходимо пользоваться какими-то косвенными методами. Трудность интерпретации результатов заключается в том, что сложно отдифференцировать, за счет чего возникает ПД нейрона — активности соматических или дендритных синапсов. Уже говорилось о том, что в области сомы существует зона НС с повышенной возбудимостью. Поэтому, естественно, если в дендритах возникает возбуждение, то, приближаясь по ним к соме клетки, оно вызовет в первую очередь деполяризацию в начальном сегменте. В этом случае при внутриклеточной регистрации будет наблюдаться медленное нарастание потенциала резко сменяемое спайком при пороге возбуждения начального сегмента. Такое медленное нарастание потенциала, отражающее возбуждение в дендритах будет существенно маскироваться ВПСП. Чтобы избежать этого, Экклс и сотрудники (Eccles et al., 1958) работали на мотонейронах кошки, которые были хроматолизированы путем перерезки передних корешков, что приводит к более четкому выявлению действия дендритных и соматических синапсов (происходит перераспределение возбудимости сомы и дендритов). Это создает благоприятные условия для выяснения особенностей возбуждения нейрона через различные синаптические входы и облегчает выяснение возможности возникновения ПД в дендритах.
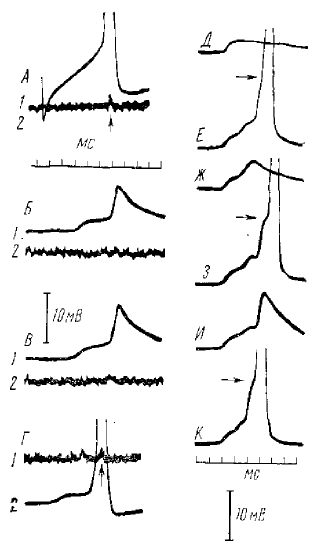
А — Г: 1 — внутриклеточные регистрации мотонейрона m. flexor digitorum longus, аксон которого был перерезан за 16 дней, 2 — регистрация от филомента, содержащего аксон мотонейрона; А — ответ на деполяризующий толчок тока; Б — Г — ответы на максимальное раздражение эфферентов группы I m. flexor digitorum longus; Д — К — ответы на афферентный залп, показывающий широкий диапазон вариабельности «частичных спайков», которые накладываются на ВПСП и вызывают полный ответ, когда деполяризация достигает критического уровня (около 13 мВ), показанною стрелками на регистрациях В, 3 и К
На рис. 1 показано, что, при постепенном увеличении афферентной стимуляции, от сомы хроматолизированных нейронов регистрируется скачкообразное увеличение пороговой деполяризации Каждый новый прирост потенциала, или, по терминологии авторов, «частичные спайки», развивается по типу «все или ни чего». Было показано, что гиперполяризация сомы не оказывает действия на частичные спайки и дает возможность предполагать, что они возникают на дендритах. Этот факт, а также возникновение ПД при разных уровнях допороговой деполяризации и отсутствие предшествующего ему ВПСП свидетельствуют, по мнению авторов, о его дендритной природе.
Более детально вопрос о генерации и проведении импульсной активности в дендритах корковых и других структур головного мозга позвоночных разрабатывался в последние годы. Особенно удобны для изучения апикальные дендриты пирамидных клеток гиппокампа и коры головного мозга, так как они имеют большую длину и параллельно ориентированы относительно друг друга, а также крупные дендриты клеток Пуркинье мозжечка. Наиболее простое строение имеет кора Аммонова рога, где пирамидные клетки расположены в один слой, а апикальные дендриты расположены перпендикулярно к этому слою.
Еще в 1955 г. Крэг и Хэмлин (Cragg, Hamlyn) показали, что при синаптической стимуляции апикальных дендритов возникает волна деполяризации длительностью 15 мс, на фоне которой может возникнуть пиковый дендритный потенциал длительностью 1—3 мс. По их мнению, этот пиковый потенциал медленно (0,4 м/с) распространяется по направлению к соме.
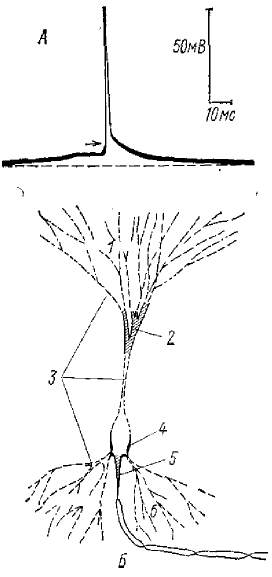
1 — апикальные дендриты, 2 — область бифуркации апикального дендрита, 3 — мембрана, характеризующаяся градуальными ответами, 4 — сома клетки, 5 — аксональный холмик, 6 — базальные дендриты
В 1961 г., изучая свойства дендритов пирамидных клеток гиппокампа, Кандель и Спенсер обнаружили (как мы уже говорили) так называемый быстрый препотенциал (БПП), который, по их мнению, представляет собой пиковый потенциал типа «все или ничего», генерируемый в пределах дендритного дерева нейрона, в достаточно ограниченном его участке, недалеко от сомы (рис. 2, А).
На рис. 2 (Б) изображена схема, где представлены соображения авторов о возможной роли БПП. Предполагается, что нейрон имеет две триггерных зоны — зону в области аксонного холмика (рис. 2, А, 5) и зону в области бифуркации апикального дендрита (рис. 2. Б, 2). БПП является ответом триггерной зоны в области ветвления (бифуркации) апикального дендрита, которая служит одновременно усилителем для более отдаленных дендритных синапсов. БПП запускают общую конечную триггерную зону в области аксонного холмика, которая в свою очередь вызовет разряд аксона и затем вовлечение сомы клетки (рис. 2, Б, 4). Благодаря этому механизму отдельные дендритные синапсы должны вызывать разряд нейрона даже тогда, когда сома гиперполяризована.
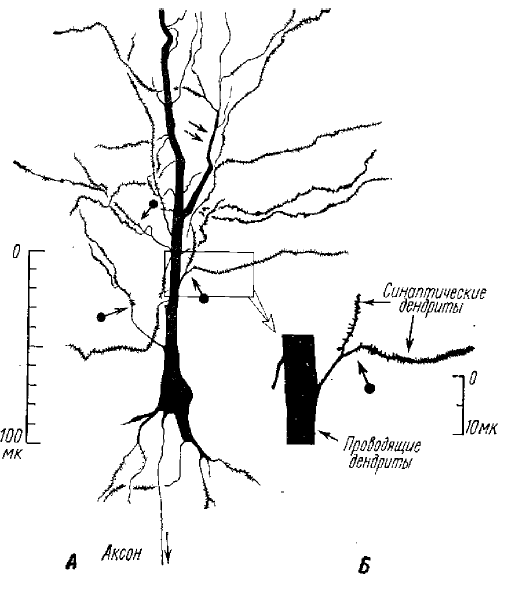
Возникновение и проведение спайка в дендритах пирамидных клеток гиппокампа изучалось также Андерсеном и Ломо (1967), В гиппокампальной коре возбуждающие синапсы расположены, как мы уже говорили, на дендритах. Андерсен и Ломо различают у пирамидных клеток гиппокампа проводящие и синаптические зоны апикального дендрита (рис. 3). Они считают, что синаптические зоны — это участки дендрита, на которых оканчиваются возбуждающие синапсы. При активации синапсов в них возникает внутрь направленный ток. В результате большой проводимости мембраны эти участки становятся шунтом для выходящего из клетки тока, что не благоприятствует возникновению спайка в синаптической области. Таким образом, по представлениям Андерсена и Ломо, синаптическая зона (субсинаптическая мембрана и близко прилегающие к ней участки поверхностной мембраны) не генерируют спайк в результате шунтирования мембраны в этом месте. Соседние области дендритов имеют более высокое мембранное сопротивление. Здесь синаптический ток создает падение напряжения достаточное, чтобы вызвать спайк. Таким образом, проводящие зоны дендрита деполяризуются выходящим током, создаваемым более дистально расположенными синапсами. В этих участках возникает пиковый потенциал, который распространяется к соме.
Интересны исследования Пурпуры и сотрудников (Пурпура, 1967; Purpura et al., 1968) относительно свойств дендритов клеток специфических таламических реле. Им изучались таламические нейроны вентробазального комплекса таламуса. Хорошо известно, что лемнисковые афференты оканчиваются преимущественно на дендритах клеток вентробазального комплекса. Однако, несмотря на это, было показано, что эти нейроны способны с большой степенью надежности следовать за сравнительно высокочастотными афферентными залпами. Проведенный анализ показал, что аксодендритиые ВПСП способны вызвать частичные или полные спайки в дендритах нейронов вентробазального ядра. Таким образом, механизм генерации спайков в дендритах может быть специальным эффективным механизмом, поддерживающим на высоком уровне отношения входа-выхода и особенно в тех нейрональных структурах, где основные афферентные проекции распределены исключительно (или преимущественно) на дендритах.
Такими афферентами являются, например, зрительные афференты tectum opticum (ТО) среднего мозга всех классов домлекопитающих позвоночных — рыб, амфибий, рептилий и птиц. Волокна ганглиозных клеток сетчатки этих животных образуют преимущественно аксодендритные синапсы с нейронами. Таким образом, большая часть зрительной информации у этих животных поступает на дендриты нейронов нервного центра. Предполагают, что дендриты нейронов ТО должны обладать сходными свойствами с таковыми нейронов вентробазального комплекса.

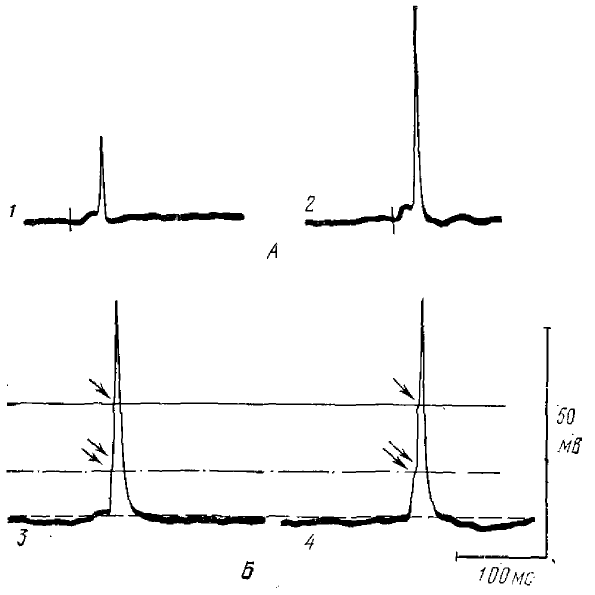
1, 2, 3 — при градуальном увеличении силы раздражения (ог 1 к 3) зрительного нерва. На вершине ВПСП возникают последовательно частичный ПД (2), а затем полный ПД (3), 4, 5 —частичяый ПД (показано стрелкой) мажет генерироваться в момент абсолютной рефрактерности сомы (4—запись фонового разряда, 5 — две последовательные реакции на раздражение зрительного нерва), 6, 7 — возникновение в ответ на раздражение зрительного нерва ПД, полного ПД и ПД промежуточной амплитуды (7), 8, 9, 10, 11 — увеличение силы искусственной гиперполяризацин нейрона (от 8 к 11) приводит к дискретным уменьшениям амплитуды ПД, 12 — гистограмма амплитуд ПД, возникающих в том же нейроне в ответ на раздражение зрительного нерва; по оси абсцисс — амплитуда в мВ, но оси ординат — число разрядов
Действительно, по данным (Гусельников, Лукьянов, 1973), дендриты некоторых нейронов ТО лягушки способны генерировать спайковые потенциалы. При внутриклеточной регистрации активности нейронов ТО наряду с полными спайками сомы клетки регистрировались небольшие пиковые потенциалы, амплитуды которых более чем на порядок отличалась от амплитуда соматического спайка (рис. 4). При градуальном увеличении силы раздражения небольшие пиковые потенциалы возникали по принципу «все или ничего», что позволяет говорить о них как о частичных спайках (рис. 4, 1, 2, 3). Искусственная поляризация мембраны нейрона слабо влияла на амплитуду частичных спайков по сравнению с влиянием на соматические спайки. Частичные спайки генерировались на заднем фронте соматического спайка, т. е. во время абсолютной рефрактерности мембраны сомы, а также в момент максимального значения следовой гиперполяризации после соматического спайка (рис. 4, 4, 5), когда генерации полного спайка не наблюдалось. Все это указывает на то, что существует зона генерации, удаленная от сомы клетки и расположенная, вероятнее всего, на дендритах. Кроме частичных и полных соматических спайков в некоторых нейронах ТО регистрировались спайки промежуточной амплитуды (рис. 4, 7). Искусственная поляризация нейрона сильнее влияла на амплитуду спайков промежуточной амплитуды, чем на частичные спайки. На рис. 4, 12 приведена гистограмма амплитуд спайков, которые возникали в одном из нейронов в ответ на раздражение зрительного нерва. Два максимума (первый и пятый) соответствуют частичным и полным спайкам, три (второй, третий и четвертый) — спайкам промежуточной амплитуды: Искусственная гиперполяризация нейрона приводила к дискретным уменьшениям амплитуды спайков (рис. 4, 8— 11). Градуальным увеличением силы гиперполяризации можно было вызвать последовательно два-три таких перехода; при этом уровни, к которым переходили амплитуды полных спайков и спайков промежуточной амплитуды, соответствовали трем первым максимумам на гистограмме. Следовательно регистрация частичных спайков и спайков промежуточной амплитуды, по-видимому, отражает блок распространения дендритного спайка в различных узлах ветвления дендритов.
Таким образом, имеется довольно много данных, свидетельствующих в пользу импульсного способа передачи возбуждения от дендритов к телам клеток в ТО лягушки.
В исследованиях Ллинаса и сотрудников (Llinas et al., 1969; Llinas, Nicholson, 1971; Nicholson, Llinas, 1971) на мозжечке рептилий (аллигатор) показана способность дендритов клеток Пуркинье генерировать и проводить спайковую активность. Отметим, что Ллинас с соавторами сделали свой вывод на весьма убедительном материале, так как ими была осуществлена внутридендритная регистрация. Специальная метка подтвердила, что кончик электрода находился непосредственно в дендрите.
Им и сотрудниками (Llinas et al., 1969) были получены данные о том, что и дендриты клеток Пуркинье мозжечка лягушки также, видимо, способны генерировать пиковую активность. Результаты внутриклеточной регистрации активности клеток Пуркинье мозжечка кошки (Martinez et al., 1971) также совпадают с этими данными. В 1968 г. Фуджита (Fujita, 1968) приходит к выводу, что клетки Пуркинье мозжечка кролика имеют несколько зон генерации пиковой активности. Способность дендритов нейронов ретикулярной формации генерировать спайк в ответ на ортодромное раздражение показал Лиманский (1964, 1965).
Что касается коры больших полушарий, то еще в 1958 г., как мы уже указывали, Бишоп высказал предположение, что в апикальных дендритах корковых клеток синаптически вызванная деполяризация может привести к возникновению затухающих «спайковых» потенциалов, которые только способствуют электротоническому распространению деполяризации на сому или прилегающий к ней участок аксона — аксонный холмик.
В работах Пурпуры (1967) была предпринята попытка выяснить способность дендритов пирамидных клеток новой коры кошки генерировать пиковые потенциалы. Им и сотр. было показано, что способностью к генерации пиковой активности обладают нейроны у кошки только в определенный период раннего постнатального развития.
В этот период пирамидные нейроны коры кошки имеют хорошо развитые апикальные дендриты и многочисленные аксодендритные синапсы. Базальные дендриты развиты слабо, а аксосоматические синапсы встречаются очень редко. Пурпура показал, что, несмотря на то что к концу третьей недели постнатального онтогенеза нейроны и синапсы новой коры кошки приобретают морфологические характеристики, сходные с таковыми взрослых животных, в дендритах пирамидных клеток на этой стадии развития возникает ПД, распространяющийся по закону «все или ничего». Однако ничего подобного нет у взрослых животных, что, как полагает Пурпура, связано с изменением возбудимости дендритов пирамидных нейронов коры. У взрослых животных лишь в некоторых случаях в неокортикальных нейронах наблюдались БПП, являющиеся отражением «частичных» спайков в дендритах. Он подчеркивает, что генерация таких «частичных» спайков дендритами нейронов новой коры исключительно редкое явление и ее нельзя рассматривать как способность дендритов взрослых животных генерировать и проводить ПД.
Чаще дендритные спайки корковых нейронов регистрируются у взрослых животных при непосредственной гиперполяризации нейрона (Воронин, 1968), эпилептиформной активности (Prince, 1969), гиперполяризации поверхности коры (Пурпура, 1967) и т. д.
1 — слабая стимуляция, 2 — сильная стимуляция, 3, 4 — два последовательных спонтанных спайка. Одиночная стрелка указывает вершину спайка аксонного сегмента, двойная — вершину дендритного спайка, прерывистая линия — уровень мембранного потенциала, штрих-пунктир — уровень возбуждения начального сегмента, непрерывная — уровень возбуждения сомы. Видно, что дендритный спайк возникает с разного уровня деполяризации мембраны нейрона
Систематические исследования обонятельного анализатора низших позвоночных показали, что способностью генерировать и проводить спайковую активность также обладают дендриты обонятельной луковицы рыб, амфибий и рептилий. На переднем фронте спайков в нейронах обонятельной луковицы выделяются два перегиба (рис. 5, Б). Один из них (обозначен одной стрелкой) выделяется при любом способе активации и является (по традиционным представлениям) вершиной спайка аксонного сегмента. Второй перегиб (обозначен двойной стрелкой) не выделяется антидромно и, вероятно, является вершиной дендритного спайка. Спайк в нейронах обонятельной луковицы не имеет стабильного уровня генерации. Это означает, что место генерации спайка удалено от микроэлектрода, а так как наиболее вероятно, что последний локализован в соме, то начальный генератор спайка следует отнести в дендриты.
Эти данные показывают, что дендриты способны генерировать пиковую активность. Латентность ответа вторичных нейронов на ортодромное раздражение на 5—15 мс больше латентности вершины афферентного ВПСП в клубочках. Следовательно, между возникновением спайка сомы и генерацией ВПСП на дендритах происходит промежуточный процесс, который, по-видимому, является распространяющимся дендритным спайком. О распространении дендритного спайка свидетельствует также зависимость его амплитуды от силы ортодромного раздражения (рис. 5, А, 1, 2). Вероятно, при слабом раздражении спайк блокируется в одном из узлов и не вызывает разряда сомы. Это может произойти в том случае, если при возрастании силы раздражения дендритный спайк растет и, вероятно, распространяется на некоторое расстояние по направлению к соме.
Таким образом, как мы видим, дендриты одних нейронов проводят пиковый потенциал, другие — нет. По-видимому, надо говорить не о свойствах дендритной мембраны вообще, а о свойствах дендритной мембраны конкретных нейронов, так как дендриты их могут иметь свои индивидуальные особенности. Не исключено также, что способ проведения возбуждения в дендритах (электротоническое, частичные спайки, распространяющиеся потенциалы действия) может изменяться от различных условий: текущей мозговой деятельности, в процессе онтогенеза, от функциональной специфики данного нервного центра и т. д.
Все сказанное показывает, что как будто бы нет сугубо принципиальных различий между свойствами мембраны дендритов и мембраной других частей нейрона. Известные к настоящему времени результаты экспериментов в общих чертах подкрепляют мнение Экклса (1959), который указывал, что пока нет оснований, «позволяющих считать, что, за исключением чисто геометрических факторов, дендриты обладают какими-то свойствами, отличающими их от сомы, от которой они отходят».1
Однако необходимо добавить, что специфику в свойствах дендритного аппарата нейронов надо искать не только в геометрических факторах, но и в особенностях строения и организации его синаптических зон, а также, видимо, в особенностях функционирования тех нервных сетей мозга, в которых данный дендрит является составляющим звеном. С другой стороны, видимо, нельзя исключать некоторых различии в свойствах мембраны сомы и дендритов и дендритной мембраны крупных и топких дендритных ветвей и в первую очередь у нейронов корковых структур, имеющих высокодифференцированные, сложные дендриты.
До сих пор дискутируется и ряд других положений о специфических свойствах дендритов и, в первую очередь, теоретические представления Грундфеста, Пурпуры и их сотрудников об электрической невозбудимости мембраны дендритов. Они полагают, что дендриты наделены лишь хеморецептивной постсинаптической мембраной, электрически невозбудимой и не способной к активному проведению по закону «все или ничего». В этом Грундфест усматривает отражение определенных эволюционных приобретений, когда дендритный аппарат, и особенно корковых нейронов, выключен из возможных электрических влияний со стороны других нервных образований и других сегментов нейрона. Изложенные факты, как мы видим, не согласуются с таким положением. По крайней мере, у толстых дендритных ветвей мембрана по своим свойствам, видимо, не отличается от остальной мембраны нейрона, она обладает электрической возбудимостью и способна проводить импульсы по закону «все или ничего». Что касается генерации и проведения спайка у дендритов многих типов нейронов, то это уже сейчас не вызывает сомнения. В то же время, если сейчас и нет прямых доказательств генерации импульсной активности в дендритах ряда нейронов и особенно сложно организованных нейронов повой коры, это еще не доказывает, что их мембрана электрически невозбудима и что при каких-то особых условиях она не будет генерировать, а возможно, и проводить спайковую активность, как это было, например, в опытах Пурпуры.
Попытка доказательства отсутствия электрической возбудимости дендритов корковых нейронов была основана также на анализе дендритного ответа коры больших полушарий на прямое ее электрическое раздражение, о котором мы говорили выше. Коснемся теперь этого вопроса подробнее. При нанесении на поверхность коры больших полушарий одиночного электрического стимула в радиусе до 6 мм можно зарегистрировать отрицательное колебание потенциала. Продолжительность потенциала до 20—30 мс. Латентный период этого ответа увеличивается в зависимости от расстояния от пункта коры, где наносится раздражение, а амплитуда зависит от силы применяемого тока. Хотя механизм происхождения этой реакции до сих пор спорен, по общепринятому мнению, она отражает ответ апикальных дендритов пирамидных нейронов коры (часто кроме дендритного ответа эту реакцию называют прямым ответом коры).
В 1951 г, Экклс предположил, что этот потенциал является постсипаптическим потенциалом дендритов и что он активно распространяется по коре посредством волокон первого слоя коры. В общем виде эта точка зрения подтверждалась фактическим материалом и других исследователей (Ройтбак, 1955, Окуджава, 1963; и др.).
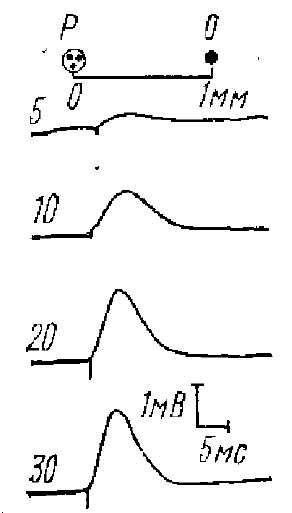
цифры па осциллограммах — интенсивность стимула в вольтах. Р — раздражающие электроды. О — отводящие электроды
Иными словами, предполагалось, что, поскольку дендриты электрически невозбудимы, электрический ток возбуждает волокна первого слоя коры, которые имеют синаптические окончания на апикальных дендритах, возбуждение’ мембраны которых происходит, таким образом, транссинаптически: возникает локальный ПСП. Для доказательства был использован большой арсенал электрофизиологических приемов исследования, которые подтверждали транссинаптическую природу этого потенциала и тем самым подкреплял и положение, об электрической невозбудимости мембраны апикальных дендритов корковых нейронов. Однако Ройтбак (1968) вновь подверг анализу механизмы возникновения этого потенциала. Он показал, что при больших силах раздражающего тока ответ коры наступает мгновенно, без заметного скрытого периода (рис. 6). Отсутствие задержки Ройтбак объясняет прямым возбуждением мембраны апикальных дендритов петлями электрического тока. Таким образом, эти данные показывают, что кроме транссинаптической природы ответа коры на прямое электрическое раздражение мембрана дендрита способна отвечать на прямое электрическое раздражение. Ройтбак предлагает термином «дендритный потенциал» обозначать постсинаптический .потенциал верхушечных дендритов при моносинантической их активации, а «прямым ответом»—ответ дендритов при больших силах тока без синаптической задержки. Эту точку зрения разделяют не все физиологи.
Другой вопрос, имеющий отношение к прямому ответу коры, более широкий и касается важного вопроса, есть ли тормозные синапсы на дендритах корковых нейронов— генерируют ли дендриты ТПСП.
Доказательство наличия ТПСП на дендритах корковых нейронов ряд исследователей пытались связать с изменениями дендритного ответа коры при ряде воздействий. По мнению Грундфеста, Пурпуры и др., кроме ВПСП дендриты генерируют и ТПСП. которые обычно маски3руются большим ВПСП, доминирование которого происходит за счет большого числа возбуждающих синапсов на дендритах. Поэтому дендритный ответ коры является электроотрицательным ответом. Иными словами, они полагают, что ответ коры является суммарным потенциалом двух типов ответов — ВПСП и ТПСП. Таким образом, если как-то искусственно подавить ВПСП на дендритах, можно было бы демаскировать ТПСП. Таким агентом, по их мнению, избирательно блокирующим деполяризующие синапсы, является гамма-аминомасляная кислота (ГАМК). При аппликации ее раствора на кору негативный потенциал при электрическом раздражении коры превращался в позитивный, т. е. демаскировалась деятельность тормозных синапсов. С точки зрения Пурпуры, Грундфеста, Окуджавы и др., все изменения амплитуды и полярности дендритного ответа коры сводятся к выключению или возбуждению возбуждающих или тормозных синапсов на верхушечных дендритах. Опытами с инъекцией ГАМК в глубь коры и в кровь (после локального разрушения гематоэнцефалического барьера) было показано также избирательное отношение ГАМК к верхушечным дендритам. У новорожденных животных ГАМК полностью блокирует дендритный ответ коры, но не вызывает извращения сто знака, что Грундфест и Пурпура объясняют поздним созреванием тормозных аксодендритных синапсов. Тот же эффект наблюдался и на примере дендритного ответа коры мозжечка, что объяснялось авторами отсутствием тормозящих синаптических окончаний на дендритах клеток Пуркинье.
Однако эти данные не подтвердили другие исследователи. Так, извращение полярности дендритного ответа коры новорожденных показано Джавришвили (1963), а ответы коры мозжечка Гольдингом, О’Лири и др. (1960). Позже были найдены и тормозные синапсы на дендритах клеток Пуркинье (Экклс, 1971). Исследования изменений дендритного ответа коры при послойной проходке ее электродом привели также к другой интерпретации извращения ответа под действием ГАМК. В опытах Джаспера, Линдсли, Мэгуна, Ройтбака и др. во время развития на поверхности коры электроположительности в глубоких слоях коры регистрировалась электроотрицательность. Поэтому авторы склонны объяснять действие ГАМК на дендритный ответ просто угнетением всех нервных элементов поверхностных слоев коры и, в первую очередь, дендритов пирамидных нейронов, а но специально только их возбуждающих синапсов. Поэтому возникновение положительного потенциала коры под действием ГАМК они рассматривают как простое дипольное отражение отрицательности глубоких слоев коры и, в первую очередь, клеточных тел и базальных дендритов пирамидных нейронов, возбуждаемых приложенным стимулом.
Это положение подкрепляется и тем фактом, что после термокоагуляции верхних слоев коры в области отводящего электрода отрицательный дендритный ответ не регистрировался, а регистрировалась электроположительность, т. е. разрушение поверхностных дендритов приводило к тому же эффекту, что и ГАМК, регистрировалось длительное дипольное отражение активности неразрушенных глубоких слоев коры. Таким образом, хотя в настоящее время нет окончательного единого мнения о ГАМК как специфическом блокаторе возбуждающих синапсов корковых нейронов, приводимые доказательства наличия тормозных синапсов на дендритах по извращению знака дендритного ответа коры сильно поколеблены.2
Итак, вопрос о наличии тормозных синапсов на дендритах корковых нейронов экспериментально не решен, хотя по аналогии с дендритами мотонейронов спинного мозга (Костюк, 1968; Kuno, Llinas, 1970) дендритами клеток Пуркинье мозжечка (Экклс, 1971; Shimono et al., 1970), латеральными дендритами маутнеровских клеток рыб (Furukawa, 1966) и дендритами митральных клеток обонятельной луковицы (Гусельникова и др., 1969, 1970), где найдены тормозные синапсы, дендриты корковых нейронов вряд ли представляют исключение. Так, например, о локализации тормозящих синапсов на дендритах корковых нейронов говорят данные Готлиба и Кована (Gottlieb, Cowan, 1972).
Из-за способности к суммации, топкой градуальности отвела и особенностей структуры и связей дендриты могут собирать информацию из многочисленных источников и подвергать ее существенной обработке. Интимные механизмы такой обработки в большинстве своем пока являются чисто умозрительными.
Для нейронов новой коры, у которых дендриты, видимо, не проводят импульсной активности, по крайней мере, ортодромно, по мнению многих исследователей, происходит топкое модулирование функции тела нейрона дендритом. Такое модулирующее влияние объясняют электрическим распространением сдвига мембранного потенциала по дендриту, который протекает по законам кабельной теории. Скорость электрического распространения потенциала зависит от емкостных характеристик мембраны; так, скорость для дендритов с диаметром 5 мкм равна 1—2 м/с. Если сопротивление мембраны больше, то электрическое проведение улучшается, возможно, за счет увеличения амплитуды ПСП в связи с повышением RBX мембраны. Однако в какую сторону меняется функция клетки при активации ее дендритного аппарата, не всегда ясно. Так, Ролл (Rail, 1964) рассчитал, что суммарное влияние дендритных синапсов более значительно из-за большой площади поверхности дендритов и большого числа расположенных на них синапсов. Поскольку преобладают возбудительные синапсы, то, по мнению подавляющего большинства авторов, локальная деполяризация дендрита должна облегчать сому и способствовать ее разряду. Однако в последние годы меняется принятая точка зрения, что торможение осуществляется только на уровне сомы. Появилось значительное количество данных о том, что торможение может иметь место и на дендритном уровне (Burke et al., 1971; Cook, Cangiano, 1972; Llinas, Nicholson, 1969; Nicholson, Llinas, 1971, и др.). Это торможение имеет, видимо, существенное значение для интегративных процессов на нейронном уровне, так как в зависимости от расположения тормозящих терминалей на дендритах они могут либо модулировать активность нейрона, либо шунтировать (выключать) отдельные афферентные входы к данному нейрону, не влияя на активность других входов. Таким образом, в этом случае дендритное торможение выступает как динамический фактор, способный изменять интегративные свойства нейрона.
Особую роль в преобразовании информации, поступающей на дендриты корковых нейронов, некоторые исследователи видят в шипиках (Chang, 1952: Lux, Globus, 1967, М. Е. Schcibe), А. В. Scheibel, 1968 п др.). Так, Люксом и Глобусом был сделан вывод, что мембрана дендритов пирамидных нейронов новой коры имеет приблизительно в 3 раза большее сопротивление, чем мембрана дендритов мотонейронов. Это сопротивление полностью определяется высоким омическим сопротивлением веточек с дендритными шипиками. Таким образом, в отличие от типа преобразования информации на синаптических контактах нейронов, не образующих шипиков, на нейронах, богатых шипиками, таких, как пирамидные клетки, можно ожидать нелинейного преобразования. Таким образом, шипики должны обеспечивать сложную интегративную деятельность дендритов. Заметное преобладание синапсов I типа на дендритах, богатых шипиками, можно рассматривать как свидетельство регулирующей роли шипиков в осуществлении суммации, управляющей активностью начального сегмента аксона. Модуляция синаптических потенциалов двумя процессами, один из которых связан с морфологией шипиков, а другой — с природой пары синаптических терминалей (возбуждающей и тормозной) подкрепляет мнение, что шипики являются пунктами сложной интегративной деятельности.
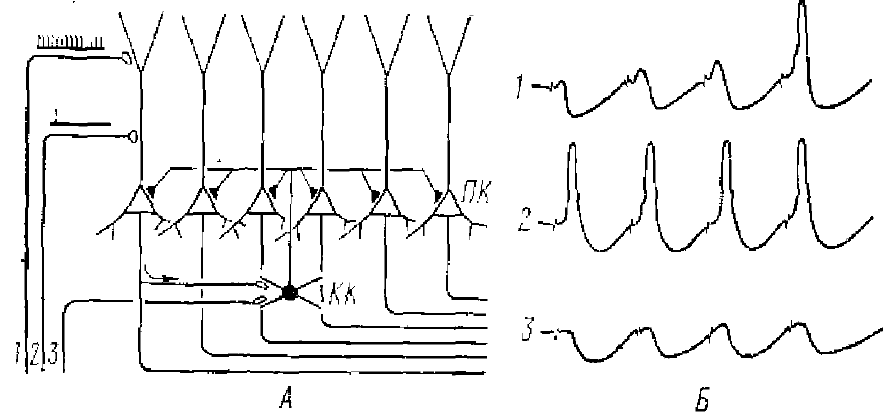
ПК — пирамидные клетки, КК — корзинчатые клетки, 1 — интеграторный путь, 2— детонаторный, 3 — путь, активирующий непосредственно КК. Схематически изображены возможности возвратного (А, 1, 2) и опережающего (1, 3) торможения
Особую роль синаптических зон на дендритах пытаются обосновать также Андерсен и Ломо (1967). Они полагают, что дендриты пирамидных клеток гиппокампа могут активироваться по двум каналам, через детонаторные и интеграторные синапсы (см. рис. 7 и 8). Детонаторные синапсы, по Андерсену и Ломо, — это синапсы на дендритах, которые возбуждают нервную клетку прямой синаптической деполяризацией триггерной зоны в области разветвления апикальных дендритов, что приводит к активности НС. Системы, в которых используются такие синапсы требуют умеренной конвергенции на конечной клетке. Такие синапсы имеют место на дендритах нейронов вентробазального комплекса таламуса, таламо-кортикальных синапсах, на пирамидных клетках соматосенсорной коры.
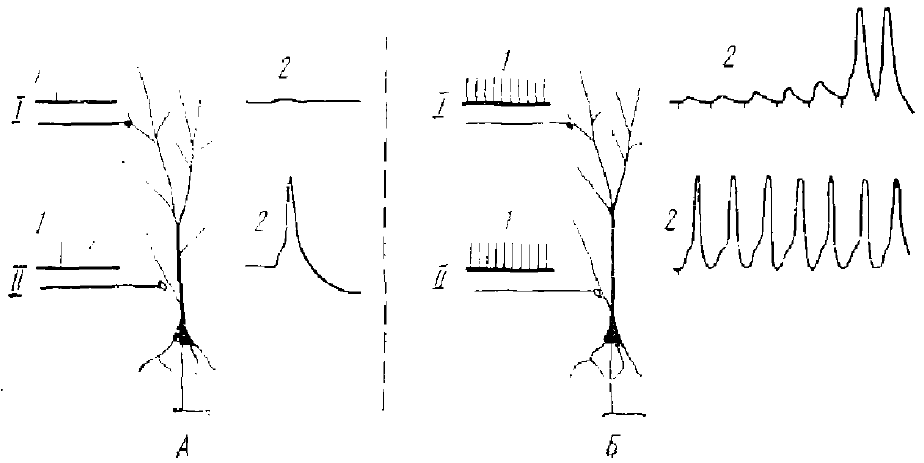
I — интеграторный канал; II— детонаторный канал, 1 — сигнал на входе, 2 — сигнал на выходе из клетки
По характеру и временным характеристикам ВПСП этих клеток можно полагать, что они возникают недалеко от сомы, в толстых ветвях дендритов. Такая детонаторная синаптическая активность должна обеспечивать значительную степень надежности и имеет высокий гарантийный фактор для распространения возбуждения по нейрональной цепи. Однако интегративные возможности такой системы значительно ниже, т. к. она обладает слабой способностью регулировать активность отдельных ее компонентов. Предполагается, что такая синаптическая активность используется в кортикальных путях, связанных с осуществлением безусловных реакций.
Примером интеграторных синапсов служат синапсы, расположенные на более тонких и конечных разветвлениях дендритов гиппокампальных и корковых пирамидных нейронов. В этом случае одиночный афферентный залп редко деполяризует начальный сегмент аксона, т. с. его деполяризация не достигает критического уровня ввиду большого расстояния от него активных дендритных зон. Для разряда нейрона требуется достаточно высокая степень конвергенции, т. е. большое количество афферентных залпов должно одновременно приходить к одному и тому же нейрону. Данные показывают, что интегративный тип синаптической активности действительно зависит от конечных ветвлений дендрита, расположенных на большом расстоянии от начального сегмента. Эффективность синаптического действия значительно возрастает при увеличении частоты стимуляции. В этом случае значительно растет амплитуда ВПСП, что Андерсен и Ломо (1967) связывают с выделением большого количества медиатора. Этот процесс они назвали частотной потенциацией.
Большую и интересную теоретическую работу, связанную со структурой дендритов и их функциональными свойствами, провела группа исследователей (Ю. А. Аршавский и др., 1966). Авторы пытались рассмотреть функциональное значение дендритов, исходя из двух допущений: 1) мембрана дендритов по своим свойствам принципиально не отличается от мембраны тела нервной клетки (обладает электрической возбудимостью и способна проводить пиковую активность), 2) механизм работы синапсов на дендритах идентичен работе таковых на соме клетки. Основным явился анализ возникновения и распространения возбуждения в разветвленных структурах. Исходя из геометрии строения дендрита, было выдвинуто положение, что «утоньшение дендритов по мере удаления от тела клетки приводят к увеличению ПСП при одном и том же синаптическом воздействии и увеличивает тем самым вероятность возникновения импульса в дендрите».3 Для доказательства этого положения авторам пришлось проделать большую работу по расчету Rвх для разных участков дендрита, так как хорошо известно, что величина постсинаптического потенциала зависит от входного сопротивления объекта, которое, в свою очередь, определяется геометрией объекта. Естественно, что по мере ветвления дендритов диаметр ветвей уменьшается и с удалением от тела нейрона RBX постсинаптической мембраны дендрита увеличивается. В результате проведенных расчетов, где ветвящиеся дендриты рассматривались как непрерывная структура, построенная из цилиндрических кабельных элементов (участок ветвления дендрита между узлами ветвления), были получены данные Rвх для разных участков.
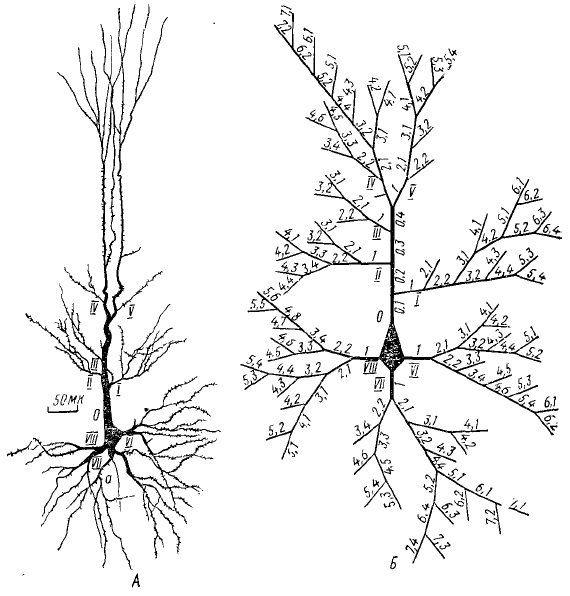
А — общий вид пирамидной клетки и нумерация главных ветвей дендритов (I, II, III, IV и т. д.); Б — нумерация мелких ветвлений дендритов (1; 2,1; 3,1 и т. д.); а — аксон
На рис. 25 схематически изображена пирамидная клетка моторной зоны коры больших полушарий кошки с нумерацией главных ветвей и тонких ветвлений дендритов, для которых авторами были проведены необходимые расчеты (RBX, электротонического затухания потенциала и т. д.).
Проведенные расчеты показали, что локальная эффективность дендритных синапсов значительно выше, чем синапсов на соме; одно и то же действие должно вызвать в конечных разветвлениях дендрита в десятки раз больший потенциал. Экспериментальное подтверждение этому можно найти в работах Фадиги и Брукхарта (Fadiga, Brookhart, 1960, 1967).
Оставалось неясным, как с удалением от тела нейрона меняется влияние синапсов на зону НС. Проведенные расчеты электротонического затухания потенциала на протяжении дендритной ветви к телу и от тела нейрона показали, что электротоническое затухание потенциала к телу клетки сильнее, что необходимо учитывать при распространении возбуждения по дендриту в анти- и ортодромном направлениях и при оценке влияния потенциалов из области дендритов на сому и наоборот. Было выяснено, что чем дальше расположен синапс на дендрите, тем меньший потенциал он вызовет в соме, несмотря на то, что исходя из расчетов в конечных разветвлениях могут возникать значительные ПСП. В связи с проведенными расчетами авторы дают ряд следствий. Так, вводится понятие неспецифического и специфического типа возбуждения нейрона. Неспецифический тип возбуждения — когда нейрон работает просто как сумматор, когда неважно, какие и где возбуждаются синапсы, а необходима лишь достаточная их активация для снижения потенциала триггерной (НС) зоны до критической величины. Естественно, что большое значение должны играть возбуждающие синапсы триггерной зоны НС.
Специфический тип возбуждения осуществляется через дендриты. Здесь уже важно не количество, а взаимное расположение возбуждающих синапсов. Так, например, активность одного или нескольких синапсов, расположенных локально на такой дендритной ветви, приводит к большому эффекту по сравнению даже с случаем, когда возбуждается большее число синапсов, по рассеянных диффузно по разным участкам нейрона.
Ю. А. Аршавский с соавторами считают, что эффективность синапсов особенно велика в самых конечных разветвлениях дендрита, где, по их образному выражению, «одной и той же спичкой легче разжечь соломинку, чем бревно». Исходя из этого, они выдвигают гипотезу спонтанной ритмической активности нейронов, полагая, что она создается за счет возвратной коллатерали от аксона пирамидной клетки к разветвлениям его апикального дендрита.
Катц и Миледи (Katz, Miledy, 1963) также полагали, что в тонких дендритных ветвях даже миниатюрные потенциалы, возникающие за счет «спонтанного» выделения квантов медиатора, могут быть надпороговыми по отношению к зоне НС, что приводит к спонтанной активности нейронов.
И, наконец, последнее, и, пожалуй, наиболее важное положение касается проведения возбуждения в дендритных узлах ветвления. По мере приближения импульса к узлу ветвления снижается фактор безопасности (гарантийный фактор), так как Rвх дендритов уменьшается по
мере приближения к телу клетки, и особенно оно падает в узлах ветвления, где может в связи с этим наступить блокада распространяющегося импульса.
Авторы замечают, что «ухудшение» и блокирование проведения в узлах ветвления может объяснить ряд описанных в литературе особенностей распространения возбуждения по дендритам: разная способность проведения в орто- и антидромном направлениях, «декрементное» проведение, крайне низкая скорость распространения возбуждения и др. Что касается последней, то, действительно, электротоническое затухание потенциала в проксимальном направлении сильнее, а скорость проведения в ортодромном направлении ниже, чем в антидромном. Возбуждение, возникшее в отдельной дендритной ветви, может блокироваться в узле ветвления, но при одновременном подходе возбуждения к узлу по двум ветвям проведение резко облегчается. Поэтому, если возбуждение, возникшее в одной дендритной ветви, не проходит через узел, то пройдет при условии возбуждения соседних ветвей, а это значит, что в узлах ветвления могут иметь место явления временной и пространственной суммации. Это и другие следствия из работы Аршавского и соавторов приводят к выводу, что разветвленная дендритная структура обладает широкими возможностями для осуществления различных логических функций.
В 1971 г. Ю. А. Аршавский с соавторами вновь вернулись к вопросу о функциональном значении структурных особенностей дендрита и пришли несколько к другим выводам о роли его узлов ветвления. Просчитав соотношение диаметров дендритных ветвей в узлах ветвления для ряда нейронов, они нашли, что в большинстве дендритных ветвей может осуществляться проведение импульса от конечных разветвлений дендрита до сомы, клетки. И лишь в области перехода импульса с дендрита на тело клетки (при ортодромном распространении) происходит гашение возбуждения. К близким данным о проведении спайка через узел ветвления в дендрите пришли Богословская и др. (1971). Однако как первые, так и последние авторы отрицают замедление проведения спайка (вплоть до его блокады) в узле ветвления. Все зависит от соотношения диаметров дендритов в узле ветвления (особенно когда дендриты образуют локальные утолщения в узле, ветвления) и интервалов между импульсами, приходящими к узлу. Это подтверждается и теоретическими расчетами (на математической модели) особенностей проведения нервного импульса через расширенный (в 3—10 раз) участок нервного волокна, проведенными Б. И. Ходоровым и др. (1970). Они также показали уменьшение амплитуды ПД, крутизны его нарастания вплоть до полной его блокады при шестикратном расширении.
Эти новые теоретические данные (как и все другие теоретические расчеты) безусловно требуют какого-то экспериментального обоснования, тем более что начальные постулаты Аршавского и сотрудников о способности мембраны дендрита проводить возбуждение но типу «все или ничего» и идентичности механизмов работы дендритных синапсов таковым сомы крайне условны и не соответствуют ряду экспериментальных данных. Кроме того, авторы использовали для основных расчетов формулу Ходжкина-Хаксли для немиэлинизированного нервного волокна, что вряд ли прямо применимо к расчетам основных свойств дендритов и особенно их конечных ветвлений,
Накопилось много литературы, посвященной попыткам не только теоретически и математически рассчитать основные принципы работы дендритного аппарата, по предприняты многочисленные усилия по созданию математических и других моделей «работающего» дендритного аппарата. Эти модели связаны в основном с вопросами распознавания и обработки сигнала на уровне дендрита (Аладжалова, 1965; Аладжалова, Шабад, 1967, 1968; Rail, 1964, 1967, 1969 и многие другие).
Таким образом, все гипотезы о функциональной роли дендритного аппарата свидетельствуют, что он, видимо, играет определенную роль в сложных процессах интеграции афферентного потока, поступающего на нейрон.
П. К. Анохиным (1973) была подвергнута критике классическая точка зрения относительно интегративных функций нейрона. Считалось, что в основе интеграции многочисленных сигнализаций, приходящих к нервной клетке, лежит сложный процесс суммации на нейроне всех субсинаптических потенциалов, результатом чего является активирование триггерных зон п генерация нервного импульса по принципу «все или ничего». Таким образом, по этой схеме интеграция сигналов происходит только на мембране нейрона, а внутриклеточной среде отводится роль пассивного проводника мембранных токов.
В частности, в лабораториях Апохина были получены весьма важные факты о существовании не только мультисенсорной, по и мультибиологической конвергенции на одном нейроне; причем возбуждения разных модельностей могут иметь и разный химизм.
П. К. Анохин полагал, что суть тонких механизмов интеграции нужно искать во взаимодействии процессов мембранного возбуждения с постсинаптической цитоплазмой и органеллами нейрона. Без этого классические представления о нейроне, созданные на основании данных электрофизиологии, вступают в противоречия с данными, полученными другими методами.
Footnotes
- Экклс Дж. Физиология нервных клеток. М., ИЛ, 1959
- Сытинский И. А. Гамма-аминомасляная кислота в деятельности нервной системы. Л., 1972.
- Аршавский Ю. А. и др. Анализ функциональных свойств дендритов в связи с их структурой. — В сб.: Модели структурнофункциональной организации некоторых биологических систем. М, 1966, с. 29.