➥ Основная статья: Тета-ритмы
Увеличение нагрузки на память
Основываясь на успехах исследований тета-ритма на животных, тета-ритм человека интенсивно начали изучать в последние два десятилетия. В одном из первых исследований, выполненных Alan Gevins (EEG systems Laboratory and SAM technology in San Francisco), изменения корковой активности при решении задач на память были исследованы с помощью электроэнцефалограмм, регистрируемых от 115 каналов. Испытуемые выполняли задания, требующие сравнения каждого нового стимула с предыдущим на основании его вербальных или пространственных признаков. В этом исследовании было получено увеличение среднелобного тета-ритма с увеличением нагрузки на память. В исследовании наблюдалось увеличение сигналов тета-ритма и улучшение выполнения задания после тренировки.
Два типа ответов в тета-диапазоне
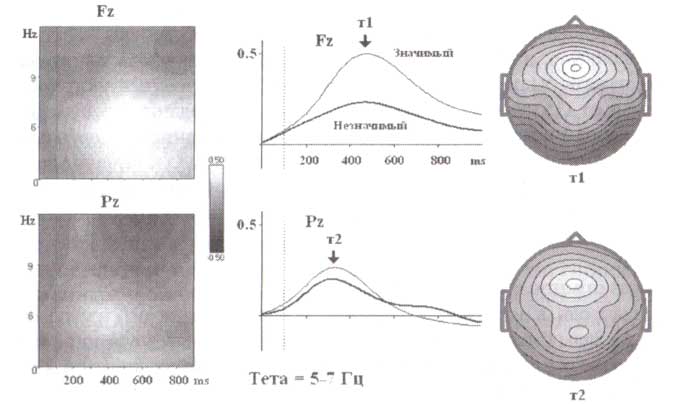
Представлены усредненные вейвлет-преобразования ЭЭГ-реакций при выполнении двухстимульной GO/NOGO-задачи. Значащие (животное на первом месте пары) и незначащие (растение на первом месте) стимулы вызывают тета подобную активность в лобных и теменных областях. Эти вызванные ритмы имеют различный временный паттерн: теменная тета-активность отличается более короткой латентностью: 350 мс по сравнению с 500 медля лобного тета-ритма. Лобная тета-активность критически зависит от поведенческого значения стимула, в то время как теменная тета-активность не зависит от значения стимула.
Ведущая роль в изучении тета-ритма людей принадлежит группе Wblfgang Klimesch из Зальцбурга (Австрия). Группа получила свидетельства того, что существует несколько типов тета-активности, по-разному соотносящихся с заданиями на память и внимание. Первый тип может быть связан с направленным вниманием или рабочей памятью. Этот тип обусловленного вниманием тета-ритма выражен в длительных тонических колебаниях в диапазоне частоты тета-ритма. Фазические вспышки тета-активности рассматриваются как основа для двух переходных типов тета-ответов: теменной тета-активности, которая связана с кодированием эпизодической памяти, и лобной активности, которая связана с извлечением из памяти. Эго заключение поддерживается многочисленными исследованиями группы Климеша, которая обнаружила, что степень синхронизации тета-ритма при кодировании определяет эффективность извлечения из памяти и что вызванная тета-синхронизация во время опознания старых слов оказывается больше, чем для новых.
Подтверждение существования двух фазических ответов в тета-диапазоне обнаруживается в работах Nobuaki Nishiyama и Yoko Yamaguchi в Brain Science Institute (BSI), RIKEN (Япония). Они записывали ЭЭГ от 64 каналов во время виртуального прохождения лабиринта. Колебания в тета-диапазоне наблюдались в виде вспышек, состоящих из нескольких циклов. Эти вспышки обычно были локализованы в двух зонах коры: лобной и теменно-височной. Первый тип вспышек был определен как среднелобный тета-ритм. Активность в теменновисочных зонах сопровождалась вспышками среднелобного тета-ритма, в то время как фазовые различия активности между этими двумя областями время от времени изменялись.
Два типа тетаподобной активности также наблюдались в исследованиях, составляющих нормативную HBI базу данных. Усредненные значения воспроизводимых ЭЭГ-ответов при выполнении двухстимульных GO/NOGO-задач представлены на рис. 1. Стимулы разделены на две категории: значащие и незначащие (Значащие стимулы — изображения животных на первом месте в парах стимулов. После предъявления этих стимулов испытуемые должны были подготовиться к следующему предъявлению стимула и нажимать кнопку, если второй стимул соответствовал первому. Незначимыми являлись изображения растений на первом месте в пробе. После предъявления этого стимула испытуемые могли расслабиться и проигнорировать всю пробу.). Можно видеть, что эти стимулы вызывают синхронизацию тета-подобной активности в лобных и теменных областях. Эти тета-подобные изменения отличаются не только в отношении пространственного распределения (лобная область в сравнении с теменной), но и в отношении временной динамики, внимания и характера привыкания к повторным пробам.
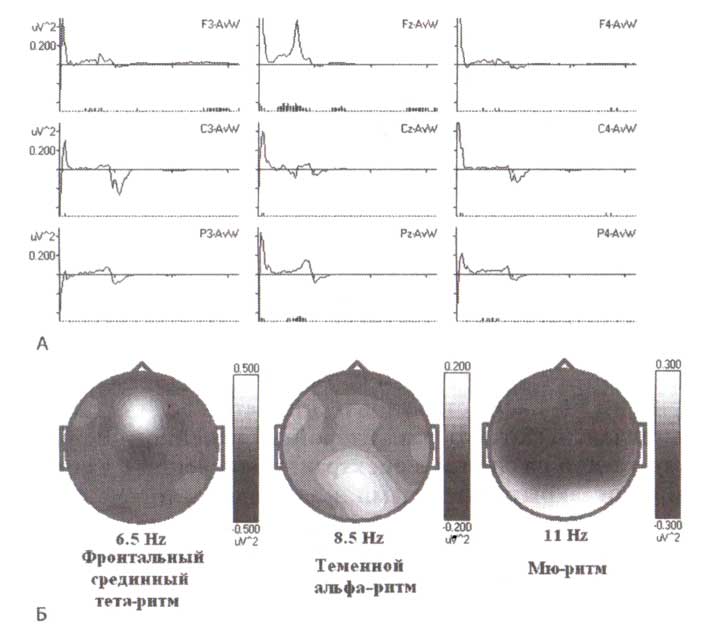
А. Результат вычитания спектров мощности, вычисленных для состояния с открытыми глазами, из спектров, полученных во время решения математической задачи для группы здоровых 13-14-летних детей. Б. Топограммы распределения разностных спектров мощности для различных частот.
Действительно, во-первых, пик латентности ответов в теменных областях меньше, чем в лобных областях. Во-вторых, лобная тета-синхронизация в сравнении с теменной тета-синхронизацией выше на значимый стимул по сравнению с ответом на незначимый. И, в-третьих, лобный тета-ритм демонстрировал привыкание к повторяющимся пробам, в то время как теменной тета-ритм этого не показал (рис. 1).
Два типа тета-подобной активности могут быть выделены простым вычитанием спектров, измеренных в состояниях с открытыми глазами, из спектров, измеренных в условиях решения задачи (в качестве примера может быть выбрано математическое задание). Результаты вычитания для группы здоровых 13—14-летних испытуемых представлены на рис. 2. Обратите внимание, что выполнение задания связано с большей мощностью ЭЭГ в тета-диапазоне (4—8 Гц) в лобной области Fz и с большей мощностью ЭЭГ в диапазоне частоты 5,5—9,5 Гц в теменной области Pz. Также обращает на себя внимание подавление мощности ЭЭГ в сенсомоторной коре из-за постоянных нажатий на кнопку, которые, как мы уже знаем, вызывают подавление мю-ритма в сенсомоторной коре.
Задачи, в которых среднелобный тета-ритм является наиболее распространенным, — те, в которых испытуемый находится как в состоянии концентрации, так и расслабления. В примерах, представленных выше, были двухстимульный GO/ NOGO-тест и математическое задание. Другие примеры заданий, связанных с индукцией среднелобного тета-ритма, — это игры, такие как тетрис, задания на вождение и др. В одном исследовании (Laukka и др., 1995) испытуемые были заняты в задаче на вождение на тренажере, в которой они должны были научиться ориентироваться и двигаться через ряд улиц, предъявляемых в виде анимации на компьютерном экране. В этом исследовании было выяснено, что процент среднелобного тета-ритма увеличился, по мере того как испытуемые учились лучше справляться с поставленной задачей и делали больше правильных решений.
Проявление в гипнозе
Состояние, которое также связывают с появлением среднелобного тета-ритма, — это гипноз. Чтобы быть загипнотизированными, люди должны сосредоточиться на себе и на человеке, который их гипнотизирует. Если они отвлечены, состояние транса будет разрушено. Высокогипнабельные люди показывают достоверно больше тета-ритма до и во время гипноза, чем низко-гипнабельные (Crawford, 1994). Другое уникальное человеческое состояние, связывающее тета-активность с вниманием, — это медитация. Есть данные о регистрации ЭЭГ во время дзен-медитаций. По мере того, как практикующие дзэн-медитации люди достигали все более и более глубоких медитативных состояний или трансов, типичные образцы активности ЭЭГ, начинаясь с альфа-активности (8—10 Гц), постепенно сдвигались в диапазон тета-активности ЭЭГ
Функциональное значение
Взаимосвязь между двумя типами тета-ответов человека и двумя типами тета-ответов у животных
Детальный анализ wavelet-преобразования нейронных ответов при выполнении различных заданий (полученных из нормативной HBI базы данных) показывает, что есть по меньшей мере два типа тета-подобной активности. Первый — среднелобный тета-ритм. Он проявляется в коротких вспышках в пределах диапазона 5—8 Гц. Этот ритм становится более выраженным с увеличением сложности задачи, оказывается выше при восприятии значащих стимулов и снижается, если тот же самый стимул представлен несколько раз. Второй — теменной альфа-тета-ритм может быть трудно различим в спектрах, вычисленных для состояний с открытыми или закрытыми глазами, и появляется только с возникновением мыслительной задачи. Теменной альфа-тета-ритм может появляться даже в виде более коротких вспышек по сравнению со среднелобным тета. В среднем этот ритм более низкой амплитуды, чем среднелобный тета, он не зависит от значения стимула и не подвержен привыканию к повторяющимся стимулам. Этот ритм не так давно был выявлен и еще не исследован подробно.
Механизмы генерации этих ритмов неизвестны. Их взаимоотношения с гиппокампальными тета-ритмами не были доказаны. Однако есть некоторые аналогии между этими человеческими ритмами и двумя типами тета-активности, обнаруженной в гиппокампальной системе в исследованиях на животных, упоминание о которых не может быть опущено в этом контексте. В исследованиях на животных два типа гиппокампальных тета-ритмов отличаются друг от друга на основе фармакологической чувствительности, частоты и поведения. Первый тип не блокируется ацетилхолиновыми антагонистами, но чувствителен к анастезирующим агентам типа уретана, этилового эфира и алкоголя, имеет частоту 6—10 Гц и возникает при произвольных движениях. Он не связан с привыканием (не адаптируется). В человеческой ЭЭГ-записи этот высокочастотный тета может быть соотнесен с теменным тета-ритмом. Он вызывается короткими вспышками, не угнетается, может быть связан с сенсорным кодированием.
Второй тип тета у животных угнетается антагонистами мускариновых (холинергических) рецепторов типа атропина. Пиковая частота этого ритма обычно 4—6 Гц (ниже, чем у первого типа). Этот тип тета-колебаний возникает при передвижениях, неподвижности и уретановой анастезии. Второй тип тета-ритма по сравнению с первым типом тренируется у интактного животного, появляется в ответ на сенсорные стимулы, когда животное находится в возбужденном состоянии. Спонтанное возникновение тета-ритма второго типа у крыс исключительно редко и имеет очень короткую продолжительность. Второй тип тета-ритма может возникать при тренировках и, вероятно, связан с научением. Возникновение второго типа тета-ритма опосредовано наличием холинергических входов в гиппокампальную формацию от центральной перегородки. В ЭЭГ человека при регистрации со скальпа аналог этого ритма может быть соотнесен со среднелобным тета-ритмом, который сильно тренируется и, вероятно, связан с эпизодической памятью.
Среднелобный тета-ритм в состоянии спокойного бодрствования может быть обнаружен в сырой ЭЭГ только в небольшой группе нормальной выборки испытуемых и соотнесен с генетически детерминированными поведенческими чертами: низким уровнем тревожности и нейротизации, высоким уровнем экстраверсии. Есть также исследования, рассматривающие среднелобный тета-ритм как признак освобождения от чувства беспокойства. Теперь известно по крайней мере три различных способа увеличения среднелобной тета-активности — медитация, биообратная связь, бинауральная техника.
Бинауральная стимуляция — важный элемент запатентованной акустической системы, разработанной Робертом А. Монро. В этом методе индивидуумы выполняют задания, включающие дыхательные упражнения, контролируемое расслабление, визуализацию и акустическую стимуляцию. Последнее задание базируется на способности мозга обнаруживать различия в частотах слуховых тонов, предъявленных в левое и правое уши. Восприятие бинаурального сигнала происходит на частоте различия между двумя слуховыми входами. Бинауральные сигналы можно легко различать в низких частотах (< 30 Гц), которые являются характерными для спектра ЭЭГ. Восприятие бинауральных тонов связано с последующим частотным ответом ЭЭГ. Это явление описано Atwater (1997).