Декларативная или эксплицитная память (англ. declarative memory or explicit memory) – это один из двух основных типов долговременной памяти человека, заключающаяся в сознательном преднамеренном воспоминании фактической информации, предыдущего опыта и концепций.
Эксплицитную память можно разделить на две категории: эпизодическую память, в которой хранятся конкретные личные переживания, и семантическую память, в которой хранится фактическая информация. Декларативная память требует постепенного обучения с многократным предъявлением стимула и реакции.
Анатомия
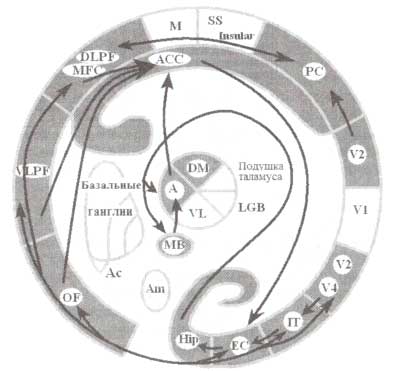
Классический случай сильной амнезии, наблюдаемый у пациента Н (одного из пациентов с билатеральным удалением медиальной височной коры), свидетельствует о важной роли этой области в обеспечении консолидации декларативной памяти. Медиальная височная кора характеризуется иерархической организацией (рис. 1): гиппокамп обладает мощными связями с энториальной корой, которая имеет сильные связи как с периренальной, так и парагиппокампальной корой; в свою очередь, эти области посылают свои проекции в область теменной и височной коры. Кроме того, гиппокамп связан с мамиллярными телами гипоталамуса и передним таламическим ядром. Известно, что повреждение этих двух структур может приводить к амнестическому синдрому, известному как корсаковский синдром. Исследования по оценке активности этих небольших близко расположенных и глубоко локализованных структур, проводимые с использованием томографии высокого разрешения (фМРТ), продемонстрировали факт их активации во время выполнения тестовых заданий на память.
Аналогичные эксперименты на животных не только воспроизводят данные, полученные при исследовании людей, но и расширяют представление о мозговом обеспечении памяти: в частности, показано, что медиальная височная кора не является единственной гомогенной структурой, вовлекаемой в обеспечение процессов формирования памяти. Иными словами, процессы, связанные с памятью, реализуются в результате сложного взаимодействия множества различных подструктур самой медиальной височной коры (рис. 1), каждая из которых вносит свой индивидуальный вклад в формирование памяти.
В соответствии с современными представлениями медиальная височная кора рассматривается как сложная система, ответственная за реализацию ряда функций, связанных с памятью, включающих кодирование, консолидацию и извлечение новых следов памяти. Важно отметить, что хранение запоминаемой информации осуществляется на уровне височной, теменной и лобной областей коры, тогда как ее извлечение реализуется при активном вовлечении гиппокампа, который всякий раз «обращается» к хранилищу информации. В конечном итоге сами по себе следы памяти становятся независимыми от структур медиальной височной коры и находятся под влиянием структур неокортекса. В частности, считается, что декларативная память на зрительные объекты реализуется на уровне вентрального пути обработки зрительной информации.
Похожие объекты, например чье-то лицо или определенная сцена, могут спонтанно запустить извлечение следа памяти. Однако в большинстве своем процессы вспоминания начинаются с целенаправленной попытки вспомнить. По всей видимости, активный процесс извлечения запоминаемой информации связан с вовлечением областей префронтальной коры. И действительно, с одной стороны, префронтальная кора получает от задних областей коры мозга в высокой степени обработанную сенсорную информацию, а также обеспечивает хранение планов действий и реализацию исполнительных функций (рабочая память, выбор, инициация и подавление действий). А с другой — префронтальная кора за счет нисходящих связей инициирует активацию запомненной информации, связанной с конкретным действием. Нейрофизиологические данные фактически подтверждают правдоподобность этой точки зрения: у пациентов с повреждениями в области левой префронтальной коры значительно нарушаются процессы произвольного вспоминания недавно запомненной информации.
Операции кодирования и извлечения
Память как единое целое проявляется в виде трех основных этапов: кодирования, хранения и извлечения. Хранение, в свою очередь, может быть долгосрочным и краткосрочным. Механизмы долгосрочного хранения исследуются клеточной биологией, изучающей структурные изменения синапсов нейронов.
Вполне логично предположить, что в целях минимизации усилий и ресурсов следы памяти кодируются и извлекаются при участии тех же областей, которые вовлекаются в обработку предъявляемых сенсорных стимулов. Первые доказательства участия сенсорных областей в обеспечении процессов извлечения из памяти были представлены в 1940—1950-х годах в работах Вилдера Пенфилда из Неврологического института Монреальского университета (W Penfield & Р. Perot, 1963). Во время открытых операций на мозге, проводимых для лечения эпилепсии, Пенфилд проводил электрические стимуляции областей затылочной и височной коры. Во время электрического воздействия испытуемые находились в сознании. Оказалось, что такие стимуляции иногда приводили к появлению воспоминаний, причем их модальность зависела от сенсорной специализации области.
Уже выполненные позже, в 1990-х годах, ПЭТ и фМРТ-исследования подтвердили эти выводы. Проведение этих исследований позволило получить прямое доказательство, что паттерны активаций метаболических процессов на разных этапах выполнения тестов на память, а именно при кодировании и извлечении информации, значительно перекрывались между собой. Так, в исследовании Вилера и др. (Wheeler et al., 2000) испытуемые запоминали зрительно предъявляемые слова в паре со звуком или изображением. Для выполнения задания необходимо было вспомнить, существовала ли смысловая связь (ассоциация) между предъявляемыми словами и сопутствующим стимулом. Такая конструкция тестового задания способствовала извлечению четких, модально-специфических следов памяти. В итоге процессы кодирования изображений ассоциировались с увеличением активности в области зрительной коры (от шпорной до веретенообразной извилины). Соответственно, кодирование слуховых стимулов вызывало активации в височной коре (от извилины Гешли до средней височной извилины). Извлечение информации об изображениях снова сопровождалось активацией в области веретенообразной извилины, а вспоминание звуков характеризовалось билатеральной активацией верхней височной извилины вблизи вторичных областей слуховой коры.
В свою очередь, операции извлечения могут быть разделены на два типа — операции сходства и воспоминания. В одной из обзорных работ Майкл Ругг и Эндрю Йонелинас (2003) представили результат сравнительного анализа данных томографических, вызванных потенциалов и нейрофизиологических исследований, в соответствии с которым два разных типа операций извлечения обеспечиваются разными мозговыми механизмами.
Нейронные модели
Cенсорная компонента события кодируется височной и теменной корой, а компонента, связанная с действием, кодируется в лобных долях. Без гиппокампа следы памяти, относящиеся к конкретным событиям, могут храниться лишь непродолжительное время за счет реверберации периодической активности в нейронных сетях этих областей.
Электрическая природа временного следа памяти обусловливает его высокую чувствительность к различным помехам (интерференции), что и лежит в основе его недолговременности. Именно на уровне гиппокампа происходит конвергенция компонент (в виде образа пространственно распределенной активности — репрезентации) в единый активационный паттерн. Образуемый таким образом след памяти характеризуется химически обусловленной долгосрочной потенциацией, индуцируемой на фоне всплесков тета-ритма. В силу этих изменений такой след может храниться достаточно долгое время. Таким образом, гиппокамп выполняет функцию временного хранения запоминаемой информации. Сохраненные следы памяти в дальнейшем могут извлекаться как сознательно при целенаправленном вспоминании, так и бессознательно, например, во время сна. Процессы извлечения обусловливают упорядочивание соответствующих следов памяти, в результате которого происходит формирование долгосрочных нейронных связей — то есть долгосрочной памяти.
Ацетилхолин как основной медиатор системы обеспечения декларативной памяти
➥ Читайте также: Нейротрансмиттеры и нейромодуляторы
Ядро перегородки как элемент восходящей холинергической системы мозга
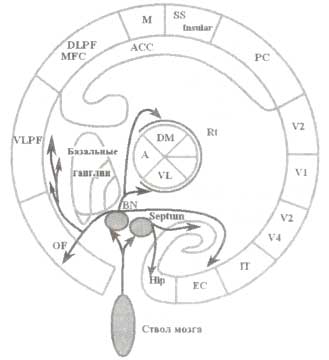
Как было выяснено выше, ключевым элементом мозговой системы эпизодической памяти является гиппокамп. Гиппокамп генерирует уникальные ритмы, называемые гиппокампальными тета-ритмами. Процесс генерации этих ритмов контролируется системами мозга, основным нейротрансмиттером которых является ацетилхолин (АХ). Ацетилхолин продуцируется в ядрах перегородки и транспортируется в гиппокамп посредством септо-гиппокампального проводящего пути (рис. 2). От интенсивности транспорта АХ зависит амплитуда осцилляций тета-активности. Следует подчеркнуть, что ядра перегородки являются продолжением холинергической системы ствола мозга и обильно иннервируются из него.
Остальные области коры (за исключением гиппокампа), включая префронтальную, медиальную височную и инсулярную кору, получают входы от другого ядра ацетилхолинергической системы — базального ядра Мейнерта. Базальное ядро преимущественно связано с лимбической корой и, следовательно, может рассматриваться как интерфейс между лимбической системой и корой больших полушарий мозга. Эфферентные связи этого ядра также направляются в область ретикулярного ядра таламуса, что обеспечивает регуляцию информационных потоков от таламуса к коре.
Известны два вида рецепторов к АХ: никотиновые и мускариновые рецепторы. Мускариновые рецепторы широко распространены по мозгу. Мускариновые рецепторы Ml-типа являются активирующими, а М2-типа соответственно тормозными. Никотиновые рецепторы локализуются преимущественно на уровне ствола мозга.
Долгосрочная потенциация и гиппокампальный тета-ритм
Теоретически в соответствии со схемой, представленной на рис. 2, ацетилхолин может модулировать процессы эпизодической памяти с помощью двух механизмов:
- увеличение амплитуды тета-ритма в гиппокампе и, соответственно, усиление долгосрочной потенциации;
- через активацию базального ядра, усиление активности многих областей коры в ответ на поведенчески значимый стимул. Множество литературных данных свидетельствует о правомерности этой схемы.
Так, фармакологические исследования показывают, что воздействия на холинергическую систему здоровых испытуемых могут приводить к существенным изменениям декларативной памяти. Например, ингибиторы ацетилхалинэстеразы улучшают декларативную память. Аналогично введение скополамина, антихолинергического агента, вызывает нарушения процессов обучения при запоминании лиц (т.е. формирование ассоциаций «лицо — имя») и сопровождается снижением уровня насыщения крови кислородом в области вентролатеральной префронтальной и нижней височной коры, а также в гиппокампе. При введении этого препарата наблюдается снижение амплитуды Р3а-компоненты , регистрируемого в ответ на предъявление редкого девиантного стимула. Существование зависимости между продукцией АХ и амплитудой Р3а-компоненты демонстрирует факт контроля холинергической системой мозга усиления нейронного ответа на новые или контекстуально девиантные стимулы.
ВП-показатели эпизодической памяти
Процессы вспоминания и эффект «старое — новое»
Существует несколько парадигм исследования нейробиологических основ эпизодической памяти. Принципиальная схема исследований универсальна — стимулы предъявляются на предварительной стадии запоминания, а непосредственно тестирование происходит во время стадии вспоминания. В исследованиях здоровых испытуемых применялись различные техники регистрации мозговой активности: фМРТ, ПЭТ и ЭЭГ. Гораздо реже применялся метод регистрации локальных потенциалов с имплантированных в структуры мозга электродов. Имплантация электродов осуществлялась исключительно в лечебно-диагностических целях.
В рамках исследовательской парадигмы «старое — новое» на этапе вспоминания показатели активности мозга регистрировались для двух видов предъявляемых стимулов. Первый тип стимула считался «старым», т.е. это стимулы, заученные испытуемым на предварительной стадии исследования. Второй тип стимула обозначался «новым», т.е. в рамках исследования такие стимулы предъявлялись впервые. Обычно наблюдаемое различие между ВП для «нового» и «старого» стимулов называется эффектом «старого — нового», а иногда — эффектом декларативной памяти. Получаемый в результате разностный ВП характеризуется выраженной височно-теменной топографией преимущественно в левом полушарии и отражает вовлечение сенсорных систем в обеспечение процессов эпизодической памяти.
Эффект «запоминания — забывания» на стадии кодирования информации
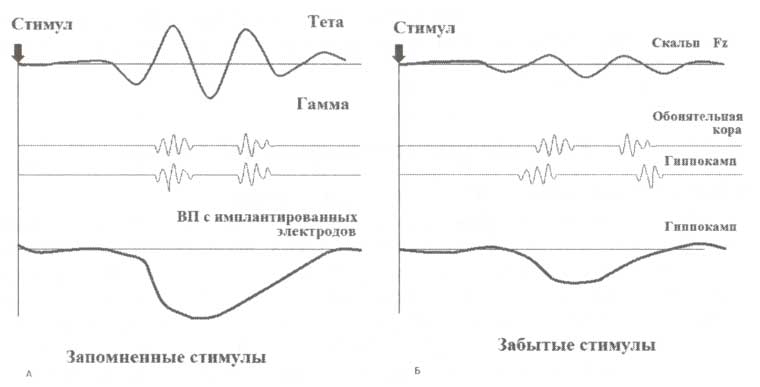
В исследовании активности мозга при процессах запоминания и забывания регистрация осуществляется на этапе кодирования стимулов, которые необходимо запомнить. В частности, параметры ВП и ЭЭГ регистрируются во время кодирования стимулов (а не на стадии вспоминания), которые впоследствии вспоминаются или забываются. Эффект наблюдаемых различий этих параметров называется эффектом «запоминания — забывания». Известно лишь несколько исследований по регистрации импульсной активности с имплантированных электродов, проводимых в рамках данной парадигмы. Результаты таких исследований схематично представлены на рис. 3. Как видно из приведенных графиков, успешное запоминание стимулов (относительно забывания) проявляется:
- как усиление ответов на уровне гиппокампа;
- увеличение когерентности между обонятельной корой (обладающей полимодальными связями с гиппокампом) и гиппокампом в гамма-диапазоне;
- увеличение амплитуды тета-ритма, регистрируемого со скальпа.