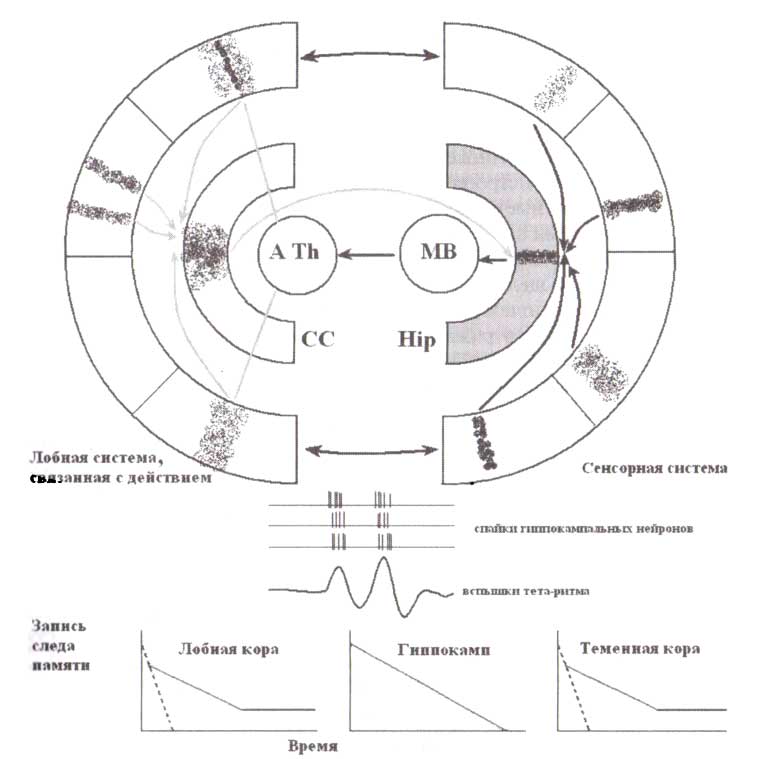
Сенсорная составляющая индивидульных эпизодов картируется в модально-специфических задних отделах коры, в то время как двигательная составляющая эпизодов картируется в префронтальных областях коры. Обе репрезентативные схемы, в свою очередь, картируются в гиппокампе. Следы, существующие за счет реверберации в префронтальных и задних отделах коры, сохраняются только в течение короткого времени (изображены прерывистой линией), в го время как следы, основанные на долгосрочной потенциации в гиппокампальных клетках, сохраняются в течение более длительного времени (обозначены серым), прежде чем они объединяются и сохраняются в долгосрочной памяти (обозначены черным).
➥ Основная статья: Тета-ритмы
Существуют данные, которые поддерживают концепцию о том, что тета-ритмы играют важную роль в обеспечении процессов памяти. Во-первых, долгосрочная потенциация (long-term potentiation, LTP) как механизм синаптической модификации, чувствительный к стадии тета-ритма, улучшается на пике цикла и ухудшается на спаде цикла (Huerta и Lisman, 1993). Во-вторых, в ряду исследований in vitro и in vivo продемонстрировано что индукция долгосрочной потенциации оптимальна, когда временной интервал между стимулами приближается к 200 мс, т.е. при частоте тета-ритма. В-третьих, разрушения перегородки не только блокируют возникновение тета-ритма в гиппокампе, но и приводят к ухудшению памяти. Эти наблюдения предполагают, что этот тета-ритм является своего рода управляющим механизмом для синаптической пластичности. Как показано, тета-ритм играет роль в нейронном кодировании места (O’Keefe и Recce, 1993). Когда крыса пересекает определенное место, гиппокампальные «клетки места» дают разряды на самой ранней стадии тета-осцилляции. Эта информация может значительно улучшить точность определения положения животного в пространстве. Другая точка зрения (Bland, Oddie, 2001) предполагает, что тета-ритм играет роль в сенсомоторной интеграции. В соответствии с этой гипотезой человеческие тета-осцилляции происходят во время эвристического, исследовательского поиска и целенаправленного поведения, а также при виртуальных движениях, когда сенсорная информация и планирование движения находятся в состоянии непрерывного изменения; по сравнению с периодами самоконтролируемого состояния неподвижности.
Следует подчеркнуть, что гиппокамп — не единственная область в мозгу, генерирующая тета-ритмы. Этот ритм был найден в большом количестве мозговых структур: парагиппокампальная кора, передняя поясная извилина, мамиллярные тела гипоталамуса, медиадорсальное ядро таламуса (рис. 1). Все эти структуры принадлежат лимбической системе. Несмотря на то что эти структуры мозга самостоятельно не способны генерировать тета-ритм, они связаны друг с другом и обеспечивают сложную нейронную сеть, выполняющую различные функции, и, следовательно, играют несколько возможных ролей для реализации тета-ритма. Gyorgy Buzaky, мировой эксперт в этой области, предложил использование термина «лимбические тета-осцилляции» вместо гиппокампального тета-ритма, что отражает сложность и распределенный характер природы возникновения тета-колебаний.
Тета-активность как отражение кванта-информации
Этому взгляду на функциональное значение лимбической системы соответствует открытие долгосрочной потенциации и ее взаимоотношений с гиппокампальным тета-ритмом. Действительно, как упоминалось выше, индукция долгосрочной потенциации оптимальна, когда стимулы повторяются с периодичностью в диапазоне тета-ритма. Чтобы вызывать долгосрочную потенциацию, ритмичное возбуждение в частоте тета-ритма не обязательно: достаточно даже двух высокочастотных разрядов с интервалом в 200 мс, чтобы вызвать долговременные изменения в синаптической работе. Эти данные явно показывают, что для того, чтобы запоминать, мозг не нуждается в постоянном тета-ритме — достаточно и двух неполных циклов. Именно поэтому цикл тета-ритма можно рассматривать как информационный квант. Этот квант, похоже, служит для функциональной связи различных лимбических структур, позволяющих кодирование некоторого эпизода нашей ежедневной жизни в воспоминание (паттерн памяти).
Гиппокамп как карта эпизодов памяти
Самое распространенное представление о функциях гиппокампа — это то, что гиппокамп кодирует в сжатой форме и временно хранит возникшие на уровне неокортекса эпизоды памяти (McNaughton, 1989). Фактически есть две части любого эпизодического события: сенсорная составляющая (включающая нейронные элементы, кодирующие различные свойства зрительных стимулов, слуховых сигналов, телесных ощущений) и связанная с действием составляющая (включающая нейронные элементы, кодирующие выполнение действия, эмоциональное состояние, побуждение). Эти две составляющие события кодируются в различных корковых областях: в париетально-височных зонах — сенсорная компонента, и в областях лобной доли — связанные с событием действия. Передняя поясная извилина служит промежуточной интегративной областью в префронтальной коре. Она интегрирует всю информацию от различных префронтальных областей и посылает результаты обработки информации к моторным областям и в гиппокамп. Хотя задняя и передняя части поясной извилины связаны друг с другом, они кодируют совершенно различные параметры события. Гиппокамп же — это зона мозга, где все различные и пространственно распределенные представления о событии сходятся в единый активационный паттерн или сегмент. С позиции анатомии гиппокамп идеально подходит для картирования. Он состоит из большого количества долей (сегментов), выровненных вдоль одной борозды. Активация ограниченного числа сегментов гиппокампа будет успешно картировать корковые представительства произошедшего эпизода в пространственный паттерн активности гиппокампа.
Тета-ритм и консолидация следов памяти
Без гиппокампа и связанных с ним структур след памяти в этих областях может быть сохранен только в рабочей памяти в форме реверберации нейронных импульсов в рекуррентных нейронных сетях. Этот след рабочей памяти существует несколько секунд и очень чувствителен к помехам — любой новый эпизод вызывает распространение следа памяти, новый след накладывается на след от предыдущего события и уничтожает этот.
Наша гипотеза состоит в том, что гиппокампальный след сохраняется на более длительное время. Продолжительность гиппокампального следа, возможно, определяется долгосрочным облегчением, вызванным высокочастотными вспышками разрядов, которые следуют за друг другом через оптимальные (в диапазоне тета-ритма) временные интервалы. Такой ход вещей позволяет гиппокампальной системе быстро воспринимать новую информацию, не разрушая ранее сформированные следы памяти, сохраненные в коре головного мозга, и в то же самое время позволяет постепенно интегрировать новую информацию с имеющейся в памяти и структурировать ее.
Вышеупомянутая модель фактически предполагает, что гиппокамп служит временным хранилищем событий. Важное событие вызывает вспышку тета-активности, которая отражает синхронную активацию гиппокампальных клеток, связанных со «сжатым» следом. Если событие не достаточно важно, то оно не вызывает вспышки тета-активности и консолидации следа не будет.
Следы в гиппокампе становятся активными или при эксплицитном воспроизведении, или при имплицидных процессах, например, во сне. Это вызывает восстановление соответствующей корковой памяти, проявляющееся в возрастании корковых взаимодействий, вероятно, вовлекающих местную синаптическую консолидацию.
Таким образом, гиппокамп использует «репрезентации» событий, чтобы неоднократно обновлять репрезентации на уровне корковых связей, до тех пор пока они не станут консолидированными и независимыми от гиппокампа. Существует много свидетельств в пользу того, что сон играет важную роль в синаптической пластичности и памяти. Было показано, что сон важен после зрительного обучения для консолидации некоторых форм процедурной памяти, например, зрительно дискриминируемых навыков. Однако облегчающий запоминание эффект сна не ограничен наблюдениями только для процедурной памяти. Так, например, известно, что лишение сна ухудшает эпизодическую память. На основе наблюдаемых явлений было сделано заключение, что эффект облегчения консолидации памяти во сне может зависеть от синхронизированной медленноволновой нейронной активности. В качестве примера — модель эпизодической памяти (Buzsaki G., 1989) постулирует, что в состоянии бодрствования информация первоначально сохраняется в САЗ-области гиппокампа, через изменения силы взаимодействий между пирамидными нейронами. Позже, во время медленного сна, синхронизированные популяционные разряды САЗ-нейронов будут «перезапускать, перепрезентовать» следы, сохраненные в САЗ-нейронной сети, и через обонятельную кору реактивировать ассоциативные корковые нейроны, воспроизводя «основные характеристики» интересующего события. В конечном счете этот перезапуск сохраненных следов памяти ведет к долгосрочным синаптическим изменениям в ассоциативных корковых сетях.
Это временное воспроизведение события (следа) в гиппокампе оказывается относительно нечувствительным к помехам, т.к. следы отдельных событий фиксируются в отдельных частях пространства гиппокампа.