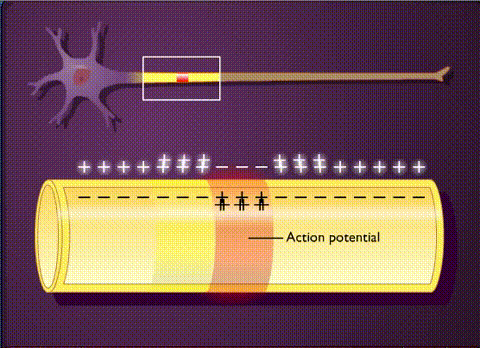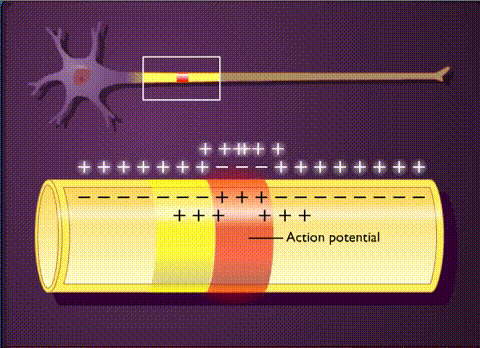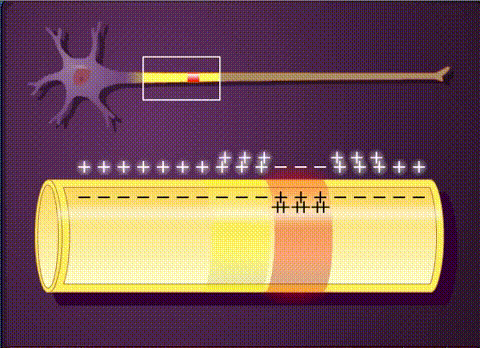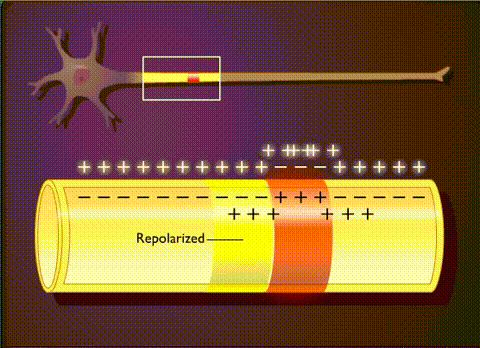
В основе электрической активности мозга лежит потенциал действия и синаптические взаимодействия между нейронами. Потенциал действия – это не что иное, как временный сдвиг (от отрицательного к положительному) мембранного потенциала нейрона (-70 мВ), вызванный потоками ионов изнутри и внутрь клетки. Он обусловлен наличием в клеточной мембране особых типов потенциалзависимых ионных каналов. Клетка срабатывает, производя потенциалы действия, который распространяются вниз по ветвям нейронов в виде цепных реакций, вызывая волну деполяризации и реполяризации. Потенциалы действия движутся только в одном направлении.
Таким образом, потенциал действия движется вдоль ветви пока не достигает конца аксона, на котором расположен синапс. Синаптическое взаимодействие может влиять на постсинаптический нейрон как тормозящим, так и возбуждающим образом. Связывание нейротрансмиттеров с рецепторами в постсинаптическом нейроне, либо перетекание заряда через щелевые контакты может вызвать либо кратковременные изменения, такие как изменения мембранного потенциала, называемые постсинаптическими потенциалами, либо более долгосрочные изменения путем активации сигнальных каскадов.
На уровне целого мозга электрическая активность проявляется как ритмические колебания электрического потенциала, наблюдаемые в нейронных ансамблях. Эти колебания, которые существуют в диапазоне частот от одного до нескольких сотен герц, доминируют в электрофизиологических измерениях крупномасштабной активности мозга и образуют мозговые ритмы и волны. Осцилляции можно охарактеризовать их частотой, амплитудой и фазой. Эти свойства сигнала могут быть извлечены при помощи регистрации электрической активности нейронов мозга с использованием частотно-временного анализа. Нейронные осцилляции и синхронизация связаны со многими когнитивными функциями, такими как передача информации, восприятие, контроль движений и память.123
Электроэнцефалография
Электрическое поле, создаваемое одиночным нейроном или совокупностью близкорасположенных нейронов, настолько мало, что не может быть зарегистрировано с поверхности головы. Электроэнцефалография или ЭЭГ отражает суммарную активность большого пула нейронов, величина которого достаточна для формирования разницы потенциала на поверхности скальпа.
Следует подчеркнуть, что электрическая активность должна быть настолько большой мощности, насколько это потребуется для прохождения сигнала через кости черепа и кожные покровы, обладающие заметным сопротивлением электрическому току. Опять-таки, исходя из приведенных необходимых и достаточных условий: активность, регистрируемая со скальпа, обусловлена относительно мощной суммарной активностью нейронального пула, — логично вытекает положение о том, что суммарная ЭЭГ в первую очередь обусловлена активностью нейронов коры как самых близко расположенных к поверхности кожи головы — месту регистрации.
Мозговые ритмы и волны
➥ Более подробно: Ритмы ЭЭГ
В центральной нервной системе (ЦНС) нейроны связываются посредством электрохимических сигналов, которые приводят к потоку ионных токов через синаптические контакты. На сетевом уровне синхронизация электрической активности нейрона вызывает ритмические колебания напряжения, распространяющиеся по областям мозга, известные как нейронные колебания или мозговые волны. Они выполняют различные физиологические функции и коррелируют с различными поведенческими состояниями. Мозг обладает множеством разных типов осцилляторов с различными периодами колебаний. Осцилляторы одновременно выдают частоты от 0,02 до 600 Гц. Классификация частотных границ позволила создать классификацию, по которой можно описывать ритмы мозга.4 На мезо- и макроуровнях эти колебания подразделяются на альфа-активность (8-12 Гц),5 которая обнаруживается в затылочной доле во время спокойного бодрствования и усиливается при открывании глаз,6 дельта (1-4 Гц), тета (4-8 Гц), мю (8-13 Гц), бета (13-30 Гц), низкая гамма (30-70 Гц) и высокая гамма (70-150 Гц) частотные диапазоны, которые могут быть измерены с помощью электроэнцефалографии.
Ритмический характер ЭЭГ определяется, по меньшей мере, тремя ритмообразующими компонентами.
- Первый – управляющее влияние ретикулярной формации среднего мозга на таламус;7
- Второй компонент – таламус и лимбическая система;
- Третий – пирамидные нейроны коры.
Предполагают, что активность коры модулируется пейсмекерами таламуса, которые в свою очередь имеют периферические входы и получают сигналы от ретикулярной формации ствола мозга.
Учеными было установлено, что целесообразной моделью для описания распределения альфа-ритма по скальпу стоит считать двухуровневую модель генерации. Первый представлен двумя базовыми генераторами в ретикулярных ядрах таламуса, второй – двумя модально-специфическими корковыми генераторами, локализованными в определенных специфичным модальности корковых областях.8
В зависимости от эмоций, чувств и состояния человека в целом всегда преобладает только одна определенная волна. В целом они привязаны к какому-либо виду мышления или ритму деятельности.
Виды мозговых волн
Альфа-волны ритмичные колебания, частотой от 8 до 14 Гц, средняя амплитуда 30-70 мкВ. Регистрируются почти у всех здоровых людей в состоянии спокойной мозговой деятельности при бодрствовании. При этом ритме организм способен производить успокаивающие нейрохимические вещества. Данный ритм выражен в теменных и лобных областях.
На данный момент ученые рассматривают несколько теории происхождения и локализации генераторов альфа-активности:
- факультативная теория пейсмекеров. Изложена П. Андерсоном и его коллегами. Она базируется на локализации центрального механизма генерации альфа ритма в таламических ядрах, влияющих на нейронную активность в соответствующих корковых областях;910
- модель корковых и таламических генераторов предложенная Ф. Лопесом да Сильва с сотр.,11 которая основана на существовании относительно независимых генераторов, находящихся как в ядрах таламуса, так и в коре;
- теория Басара Е.12 утверждающая наличие множества мультифункциональных селективно распределенных по мозгу генераторов альфа- ритма, образующих диффузно-распределенную альфа-систему;
- В свою очередь, Williamson S.I. полагает, что в мозговой коре есть самостоятельные автогенераторы, т.е. дискретные источники осцилляций, называя такие источники альфа диапазона «альфонами» . Кроме того, есть предположение, что подобным же образом в коре головного мозга существуют и генераторы бета-, тета-. дельта- ритмов.13
В последних исследованиях были выделены новые пограничные подгруппы альфа-волн – Мю волны (Тальфа) в диапазоне от 7 до 13 гц. Активизируются при визуализации движений или при физ. активности. Исходят из передних частей полушарий.
Бета-волны – высокочастотные (13-100 Гц), но низкоамплитудные (14-40 мкВ). Их регистрируют в лобных областях в нормальном состоянии бодрствования, мы внимательны и сконцентрированы. При взволнованном или тревожном состоянии генерируются быстрые бета-волны, а при ощущении угрозы и опасности – сверхбыстрые.
Гамма-волны (разновидность бета-ритма) – самые быстрые волны. Находятся в диапазоне 30-170 Гц (в некоторых источниках до 500 Гц). Амплитуда крайне низка – ниже 10 мкВ. Встречаются при решении сложных проблем или задач, описывается состояние «эврики». Так же возможны появления при волнении и страхе.
Тета-волны – медленные волны частотой 4-8 Гц и амплитудой 20-60 мкВ. Эти волны наиболее интенсивны в затылочной и височных областях. В норме регистрируются у детей или при эмоциональном стрессе у взрослых (в подавленном состоянии, фрустрации).
Дельта-волны – все низкочастотные волны на ЭЭГ (ниже 3,9 Гц). Низкоамплитудные (20-30 мкВ). Генерируются при глубоком сне или при коме. У детей могут проявляться при серьёзных органических поражениях мозга. Они считаются «истинно корковыми» волнами из-за их автономности при активности нижележащих областей мозга.
Ритмы головного мозга
Альфа-ритм
➥ Более подробно: Альфа-ритм
Регистрируется преимущественно в затылочных отведениях в виде синусоидального, модулированного в веретена по амплитуде ритма частотой 8-13 Гц, амплитудой до 100 мкВ. По направлению к передним отведениям его амплитуда и индекс уменьшаются. Наибольшая амплитуда альфа-ритма отмечается в состоянии спокойного, расслабленного бодрствования, в темном помещении, при закрытых глазах. При открывании глаз он редуцируется, а при закрывании глаз восстанавливается (рис. 1-3).
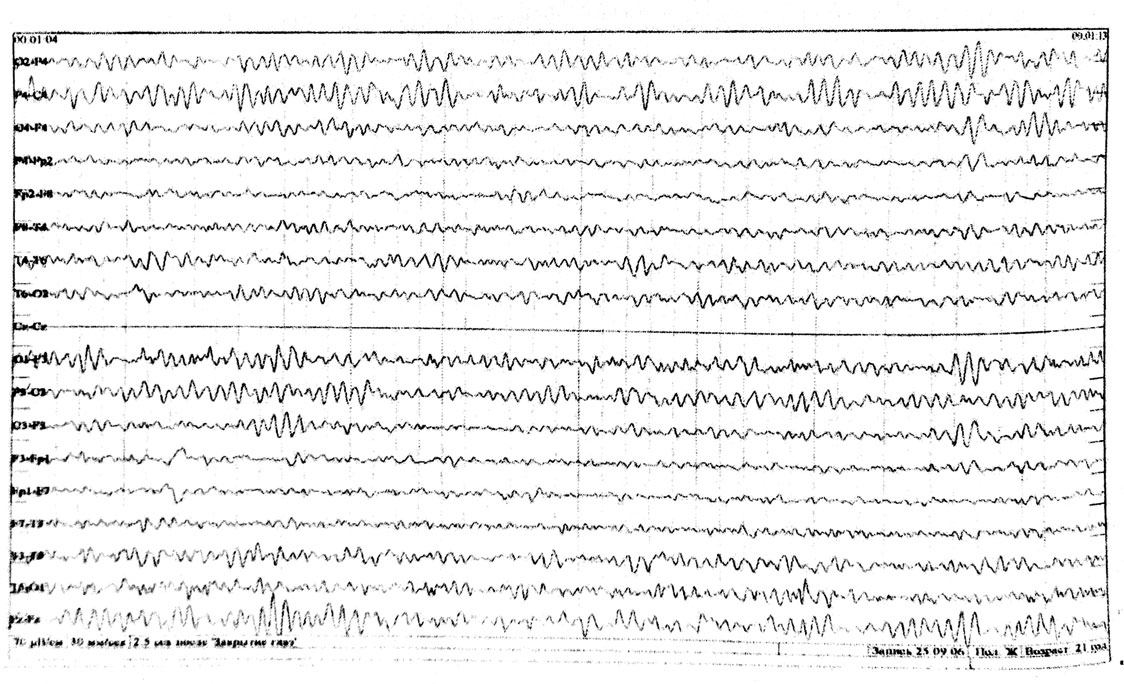
Сенсомоторный мю-ритм
➥ Более подробно: Мю-ритмы
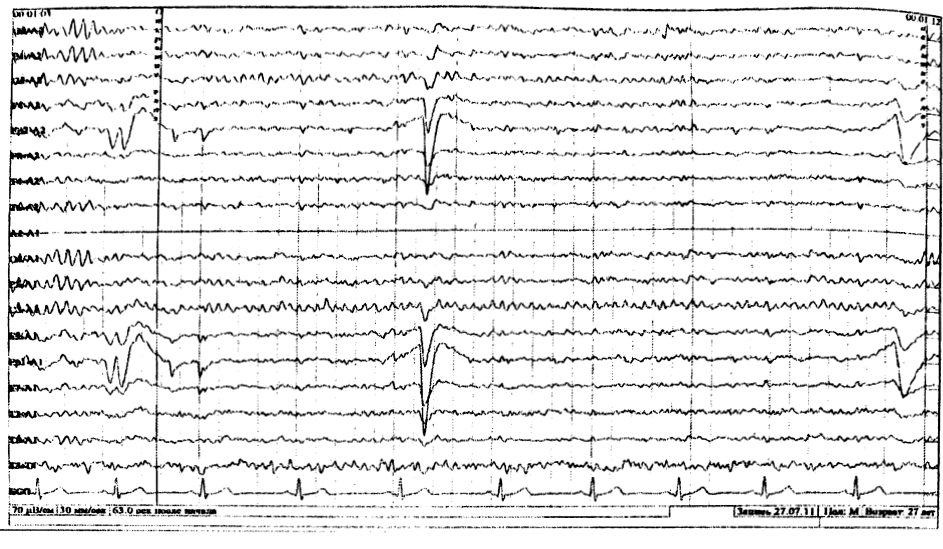
Встречается преимущественно в молодом возрасте. Выявляется на ЭЭГ в центральных отведениях в виде аркообразного ритма, частотой равной или немного превышающей альфа – ритм (рис. 4).
На открывание и закрывание глаз не реагирует. Активируется умственной нагрузкой и психическим напряжением. Редуцируется при активации двигательной активности в конечностях (проба со сжатием кулаков).
Бета-ритм
➥ Более подробно: Бета-ритмы
Регистрируется преимущественно в передних отведениях, частотой 14-40 Гц, амплитудой до 20 мкВ , шумоподобный, не реагирует на открывание и закрывание глаз (рис. 5).
Бета – ритм связан с соматическими, сенсорными и двигательными корковыми механизмами и дает реакцию на двигательную активность или тактильную стимуляцию. При выполнении или умственном представлении движения бета-ритм исчезает в зоне соответствующей корковой проекции.
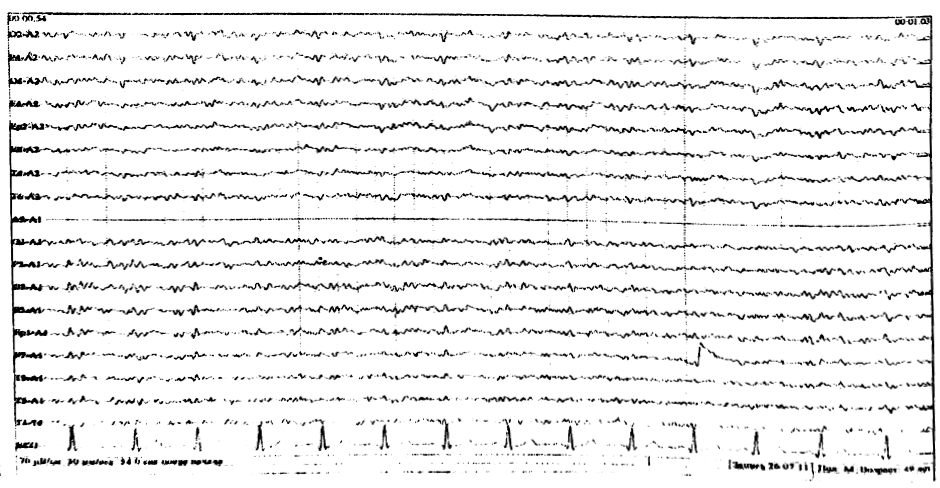
Тета – колебания
➥ Более подробно: Тета-ритмы
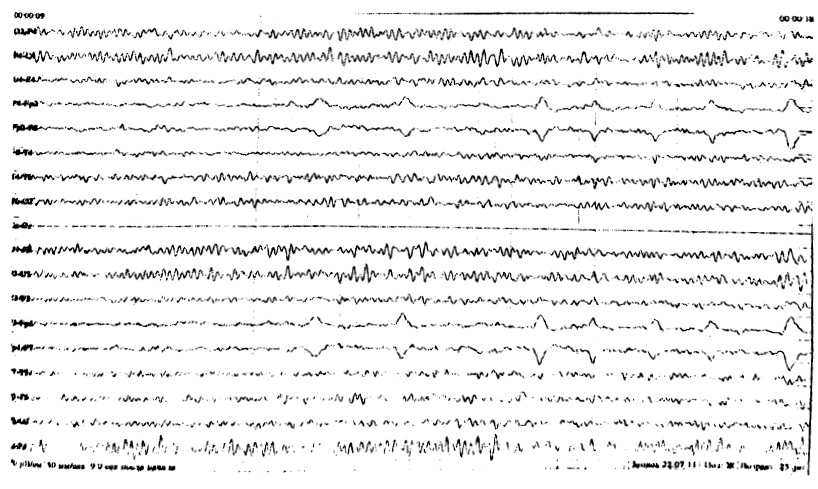
Регистрируются преимущественно в передних отведениях, в виде единичных волн, по амплитуде не превышающих основной корковый альфа-ритм (рис. 6). В отдельно-взятой эпохе ЭЭГ количество тета – волн должно составлять у молодых до 20%, постепенно увеличиваясь с возрастом до 40%.
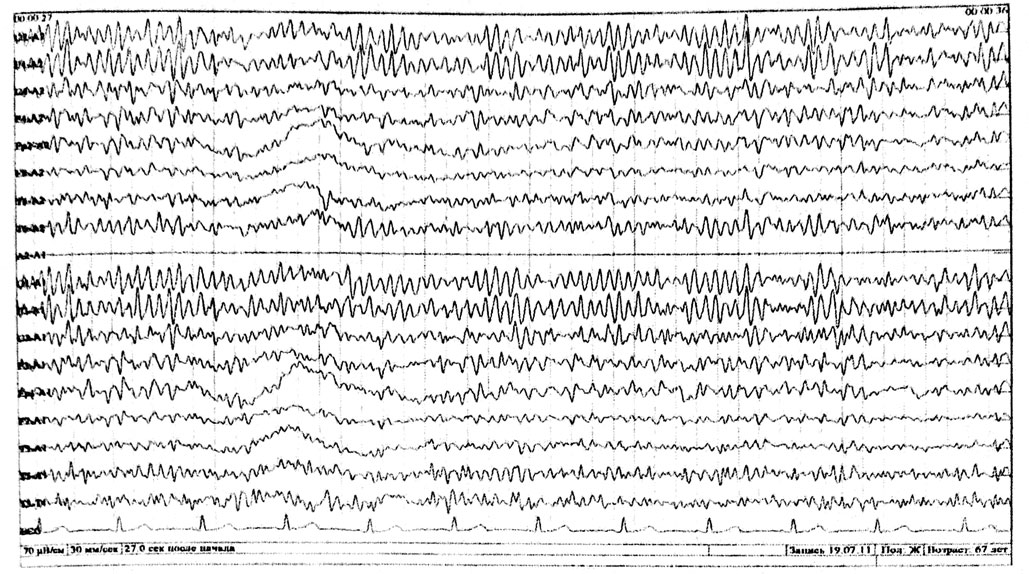
Нормальная ЭЭГ взрослого бодрствующего человека
➥ Более подробно: Нормальная ЭЭГ бодрствования
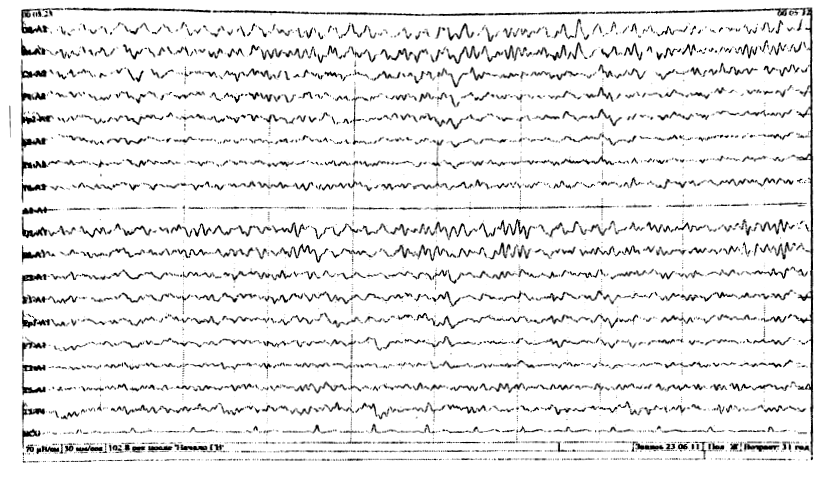
У большинства здоровых взрослых в состоянии спокойного бодрствования на ЭЭГ доминирует альфа-ритм, с амплитудным преобладанием в теменнозатылочных отведениях. По направлению к передним (лобно-центральным отделам) альфа-ритм убывает по индексу и амплитуде и сочетается с бета активностью (рис. 6). У 10-15 % здоровых обследуемых, регулярный альфа-ритм не превышает 20 мкВ и по всему мозгу регистрируются высокочастотные, низкоамплитудные колебания, которые свидетельствуют о преобладании в мозге десинхронизирующих влияний. Это тип ЭЭГ связан с аутосомнодоминантным геном. Л. Р. Зенков указывает и на половые различия ЭЭГ: женщинам свойственны более высокие частоты альфа – ритма и большее количество бета-активности. Автор делает вывод, что эти данные определенным образом коррелируют с половыми различиями психологических характеристик, свидетельствуют о более высоком уровне активации у женщин, имеют генетическую обусловленность и могут быть связаны с гормональными особенностями. У некоторых здоровых обследуемых отсутствует нормальный альфа-ритм, а вместо него регистрируется активность частотой 14-18 Гц, амплитудой около 50 мкВ, с преобладанием в затылочных отведениях и снижением амплитуды по направлению к передним отделам (рис.2). При умственной нагрузке, эмоциональном напряжении этот ритм исчезает (реакция активации).
Альфа-ритм выражен в затылочных отведениях, модулирован по амплитуде, частотой 9 Гц. Межполушарная асимметрия отсутствует. Зональные различия сохранены. Бета-ритм преобладает в передних отведениях, невысокой амплитуды легко деформирован тета- волнами. Медленная активность представлена единичными тета- волнами, по амплитуде не превышающими фон, регионарно преобладающих в передних отведениях, без склонности к группированию и фокального акцента. Дельта волны – отсутствуют.
Читайте также
- Пачка спайков / вспышка (Bursting)
- Нейронная активность
- Модель одиночного диполя
- Активность одиночного нейрона
- Активность нейронной популяции
- Локальные потенциалы
- Потенциал действия нейрона
- Вызванные потенциалы
Footnotes
- Fries P (2005). “A mechanism for cognitive dynamics: neuronal communication through neuronal coherence”. Trends in Cognitive Sciences. 9 (10): 474–480. doi:10.1016/j.tics.2005.08.011.
- Fell J, Axmacher N (2011). “The role of phase synchronization in memory processes”. Nature Reviews Neuroscience. 12 (2): 105–118. doi:10.1038/nrn2979.
- Schnitzler A, Gross J (2005). “Normal and pathological oscillatory communication in the brain”. Nature Reviews Neuroscience. 6 (4): 285–296. doi:10.1038/nrn1650.
- Buzsáki, G (2006). The Rhythms of the Brain. Oxford Press.
- Foster, JJ; et al. (July 2017). “Alpha-Band Oscillations Enable Spatially and Temporally Resolved Tracking of Covert Spatial Attention”. Psychological Science. 28 (7): 929–941. doi:10.1177/0956797617699167.
- Berger H, Gray CM (1929). “Uber das Elektroenkephalogramm des Menschen”. Arch Psychiat Nervenkr. 87: 527–570. doi:10.1007/BF01797193.
- Фокина Ю. О., Павленко В. Б., Куличенко А. М.. “ВЕРОЯТНЫЕ МЕХАНИЗМЫ ГЕНЕРАЦИИ ЭЛЕКТРОЭНЦЕФАЛОГРАММЫ”// Ученые записки Крымского федерального университета имени В. И. Вернадского. Биология. Химия, vol. 20, № 4 (59), 2007, pp. 96-108
- Исаичев C.A.. Осипова Д.С.. Коптелов Ю.М. Дипольные модели генераторов альфа-ритма // ЖВНД. – 2003. – Т. 53. № 5. – С. 577-586
- Andersen P.. Andersson S.A. Thalamic origin of cortical rhythmic activity / Handbook of Electroencephalography and Clinical Neurophysiology / Ed. Amsterdam: Elsevier. 1974. – Vol. 2 , – P. 90
- Andersson S. A.. Holmgren E. Theoretical consideration on the synchronization of thalamo-cortical activity Subcortical Mechanisms and Sensorimotor Activities / Ed. Frigyesi T.L. Bern: Huber. 1975. – P. 229-250.
- Lopes da Silva F.H.. van Lierop T.H.M.T.. Schrijer C.F.M.. Storm van Leeuwen W. Essential differences between alpha rhythm and barbiturate spindles: spectra and thalamo-cortical coherence Electroenceph. clin. – 1973b. – V. 35. – P. 626-639
- Basar E. Brain function and oscillations. – Berlin. New York: Springer. 1998. – 467 p
- Лобасюк Б.А. Роль ретикулярной формации ствола мозга в механизмах электрогенеза Нейрофизиология. – 2005. – Т. 37. №1. – С.39-45