Реакция перестройки (РП) или навязывания ритма заключается в изменении спонтанной активности коры мозга под влиянием ритмических световых раздражений. В результате такой перестройки электрическая активность коры мозга приобретает ритмический характер с частотой либо равной, либо в целое число большей или меньшей, чем частота ритмических световых раздражений.
Относительно диапазона частот, в котором может возникать РП, имеются разноречивые высказывания. Так, Эдриан и Мэтьюз (Adrian, Mattws, 1934) полагали, что потенциалы коры мозга могут следовать за ритмом световых раздражений до 20—30 имп/с. По Джасперу (Jasper, 1937), верхняя граница частот РП у человека достигает 55—60 Гц, по Тири (Thiry, 1951) — 70 Гц, по Манди-Кастлу (Mandy-Castle, 1953) — 44 Гц. Разноречивые данные получены и для животных. Такой разнобой в экспериментальных данных, видимо, во многом зависит от индивидуальных свойств испытуемых и от характера фоновой электрической активности коры. У лиц с хорошо выраженным фоновым α-ритмом частотный диапазон РП, как правило, уже, чем у лиц с плохо выраженным α-ритмом.
Характеристики РП зависят от методики проведения экспериментов, и прежде всего от параметров применяемого ритмического светового раздражения. Так, в работах ряда исследователей было показано, что увеличение яркости светового раздражения приводит к увеличению амплитуды электрических колебаний, составляющих РП, и расширяет ее частотный диапазон. Однако увеличение интенсивности раздражения сверх некоторой критической ведет к ослаблению реакции. Подобные данные получены и на человеке. Показано, что существует оптимальная интенсивность мелькающего света, при которой РП имеет максимальную амплитуду и наиболее широкий частотный диапазон. Найдена зависимость характеристик РП и от других параметров раздражения — степени контраста, соотношения периодов света и темноты (Ильянок, 1960, 1961).
Противоречивые данные о частотном диапазоне РП в значительной степени могут быть связаны с зависимостью характеристик реакции от параметров светового раздражения, поскольку нет стандартной методики провокации этой реакции, которая применялась бы большинством исследователей. Такого рода разногласия могут объясняться и произвольностью оценки наличия или отсутствия реакции при крайних параметрах раздражения. Амплитуда потенциалов РП тем меньше, чем больше отличается частота ритмического светового раздражения от оптимальной. Предельно высокая частота, при которой еще удается обнаружить реакцию, при визуальной оценке будет существенно зависеть от субъективного фактора, а при объективных методах оценки от того, насколько совершенна методика обнаружения реакций с малой амплитудой. Например, применение остронастроенных фильтров, выделяющих из электрограммы частотные компоненты, соответствующие частоте применяемого раздражения, позволило выявить ритмическую реакцию на мелькающий свет при частоте мельканий свыше 100 в 1 с, а иногда даже до 160 (Ильянок, 1959, 1960), тогда как в более ранних работах других авторов не удавалось обнаружить РП и при частотах, вдвое меньших.
Что касается локализации РП в коре, то отмечена приуроченность ее к задней части полушарий, преимущественно к затылочной области. В теменной области РП появлялась нерегулярно и имела небольшую амплитуду. К выводу о строгой локализации РП в затылочной области пришли Гасто и Хантер (Gastaut, Hunter, 1950), т. к, в других областях коры она регистрируется только при искусственном повышении возбудимости путем введения метразола. Однако большинство авторов считает, что РП не ограничена проекционной зоной зрительного анализатора и может быть зарегистрирована и в норме в соседних областях коры, хотя в них она выражена слабее. Можно наблюдать РП на ритмический световой раздражитель даже в передних областях мозга человека (Walker et al., 1944; V. Walter, W. Walter, 1949; Зислина, 1955; Копылов, 1957) и животных (Ливанов, 1940, 1944; Gastaut et al., 1958, 1959 и др.) Но амплитуда РП здесь невелика и фокус максимальной амплитуды и реакции четко локализуется в затылочной коре.
Электрические потенциалы, изоритмичные, с прерывистым световым раздражением, могут наблюдаться не только в коре, по и в подкорковых образованиях: в ретикулярной формации среднего мозга, латеральных ядрах таламуса, красном ядре, в вентромедиальном и переднем и задне-медиальном таламусе.
Важным для понимания природы РП является наличие оптимальной частоты ритмических световых раздражений, при которой РП возникает легче всего, имеет наибольшую регулярность н амплитуду. Эта частота близка к частоте основного ритма ЭЭГ — α-ритма человека или аналогичных ему ритмов у животных. По данным одних авторов, эта оптимальная частота световых мельканий практически совпадает с частотой фонового α-ритма или α-подобного ритма, по другим данным, она несколько превышает частоту основного фонового ритма. Это может служить указанием на то, что по своей природе РП близка к спонтанной фоновой активности типа α-ритма и может быть той же самой α-активностью, провоцируемой ритмическими. раздражениями зрительного анализатора (подобно тому как реакции вовлечения и усиления можно считать веретенообразной активностью, провоцируемой ритмическими раздражениями таламических ядер).
О близком сходстве РП со спонтанной α-активностью говорят и другие свойства, в первую очередь — локализация их в коре. Для обеих реакций характерна максимальная выраженность в затылочной коре и довольно широкое распространение (при более слабой выраженности) на другие области коры.
Этих косвенных признаков безусловно недостаточно для заключения об образовании РП. Хотя ряд авторов придерживается именно такой точки зрения, она отнюдь не является общепринятой. Существует мнение в противоположного характера, согласно которому РП является ритмической последовательностью потенциалов, возникающих в ответ на каждый световой стимул, которые по своей природе могут значительно отличаться от α-активности.
Отметим, что электрические реакции на ритмические световые раздражения, регистрируемые у наркотизированных животных, у которых α-подобная активность резко угнетена, действительно едва ли имеют что-либо общее с α-подобной активностью. В таких случаях часто можно видеть, что электрограмма состоит из отдельных колебаний типа вызванных потенциалов (Bartley, 1936, и др.). Однако вряд ли следует реакцию такого рода относить к категории РП ритма, наблюдаемой у бодрствующего человека и животных.
Нашли (см. Гусельников, Супин, 1968), что эти два типа реакций на ритмические световые раздражения имеют совершенно различные механизмы. Если реакция коры на ритмическое световое раздражение является последовательностью отдельных вызванных потенциалов, то они имеют, как правило, величину не на много большую, а в большинстве случаев меньшую, чем при редких (одиночных) таких же раздражениях. Классическая же РП имеет значительную амплитуду только при ритмических раздражениях, частота которых не слишком отличается от оптимальной (частота α-активности, или α-подобной активности), т. е. существует не только верхняя, но и нижняя граница частотного диапазона РП. У человека эта нижняя граница лежит около 5 Гц. Разумеется, вызванные потенциалы на редкие (одиночные) световые раздражения возникают и у человека и могут быть зарегистрированы в затылочной коре, но они имеют очень небольшую амплитуду, как правило, во много раз меньшую, чем амплитуда волн РП на оптимальные частоты.
Но и в отношении таких действительно ритмических реакций, как РП в ЭЭГ человека и вполне сходные с ней формы реакции у животных, существуют мнения о формировании их на основе последовательности вызванных потенциалов, которые в принципе аналогичны потенциалам, возникающим при одиночных раздражениях, и отличаются от них только вследствие того, что при ритмических раздражениях заметно сказывается влияние предыдущих ответов на последующие.
Так, ряд авторов (Gastaut, Hunter, 1950; Ciganek, 1958, и др.) пришли к выводу о формировании РП на основе вызванных потенциалов типа первичных ответов (ПО), прослеживая постепенные изменения электрических реакций коры на световые раздражения по мере увеличения частоты стимулов от очень редких (одиночных) до частых ритмических. В этом случае при редких вспышках света возникает сложная реакция на каждый стимул, состоящая из ПО, вторичных вызванных потенциалов и ритмического разряда последействия в виде нескольких волн синусоидальной формы. По мере уменьшения интервалов между стимулами происходит постепенное «отсечение» более поздних компонентов ответа, так что, по мнению авторов, при достаточно высоких частотах раздражения остаются только самые ранние компоненты реакции — ПО на каждый стимул.
Гасто и Хантер (Gastaut, Hunter, 1950) считают возможным сделать даже более определенный вывод, рассматривая РП на достаточно высоких частотах последовательностью лишь первых (положительных) фаз ПО. Цыганок (Ciganek, 1950), однако, полагает, что как раз при высоких частотах раздражения реакция, складывающаяся из вызванных потенциалов, исчезает и заменяется реакцией какой-то другой природы, состоящей из колебаний потенциала синусоидальной формы.
Бесспорно, что простое визуальное прослеживание изменения электрических реакций коры при постепенном увеличении частоты световых мельканий имеет, как метод выяснения природы РП, значительные недостатки. Наблюдающееся «отсечение» поздних компонентов реакции, наступающее в том случае, когда интервал между стимулами становится короче латентного периода данного компонента, не обязательно свидетельствует об исчезновении этих компонентов. Они могут по-прежнему существовать, накладываясь на последующие ответы. Конечно, такое наложение должно изменять форму ответов. Но, поскольку изменение частоты раздражения само влияет на форму вызванных потенциалов, трудно обнаружить, имеется ли одновременно и искажение их формы за счет наложения поздних компонентов предыдущих ответов. В таких условиях трудно установись, сохраняются ли при учащения раздражений те же ответы, которые наблюдаются при редких стимулах, и только форма их несколько изменяется, или же эти ответы постепенно замещаются потенциалами совершенно иной природы, в том числе и потенциалами, аналогичными по природе фоновой активности. Визуальная оценка совершенно недостаточна для решения этого вопроса, и работы с прослеживанием изменений формы реакции при разных частотах раздражения сами по себе не могут дать надежных данных относительно состава и механизмов РП.
Таким образом, рассмотренные до сих пор работы не дают точного ответа на вопрос о том, в каких отношениях находятся спонтанная α-активность (или α-подобная активность) и РП. Ряд свойств РП указывает на то, что она может являться искусственно вызываемой и модифицируемой α-активностью.
Нами (Гусельников, Супин, 1962, 1968; Супин, 1963) была сделана попытка более детального экспериментального анализа механизмов РП, в частности, с целью проверки предположения об образовании РП на базе α-активности (или α-подобной активности). В качестве основной. экспериментальной модели были выбраны реакции, наблюдаемые в электрической активности затылочной коры бодрого, ненаркотизированного кролика. В затылочной коре бодрого кролика наблюдается спонтанная ритмическая активность, которая может быть одним из наиболее вероятных аналогов α-активности человека (α-подобная активность). Если считать это положение справедливым, то затылочную кору бодрого кролика следует признать адекватной экспериментальной моделью и для решения вопросов, связанных с вызванной ритмической активностью α-подобного типа, т. е. РРП (ритмического разряда последействия).
➥ Более подробно: Сигма (σ)-ритм или веретенообразная активность. Реакция вовлечения реакция усиления
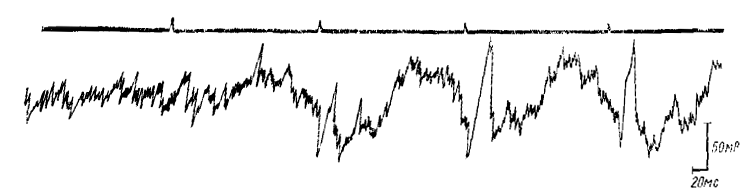
Рис. 1. Формулирование реакции перестройки ритма в затылочной коре ненаркотизированного кролика в ответ на ритмические световые вспышки с частотой около 5 в 1 с: отметка раздражителя вверху (по Гусельникову и Супину, 1962)
При ритмическом световом раздражении волны РРП, вызываемого каждым световым стимулом, увеличиваются по амплитуде от стимула к стимулу, если частота световых вспышек близка к собственной частоте РРП (около 5 в 1 с). Если два одинаковых световых стимула даются с интервалом около 200 мс, РРП, вызванный вторым стимулом, имеет, как правило, большую амплитуду и большее количество волн, чем РРП на одиночное раздражение. Еще более отчетливо увеличение амплитуды волн РРП наблюдается при ритмическом световом раздражении с частотой около 5 имп/с. Нарастание амплитуды волн происходит в течение первых двух—четырех стимулов и быстро достигает максимума. Таким путем формируются непрерывные ритмические колебания потенциала, состоящие из медленных (длительностью около 200 мс каждая) волн, идентичных первой волне РРП на одиночное раздражение, но большей амплитуды (рис. 1). Таким образом, основной рисунок биопотенциалов при частотах раздражения порядка 3—7 имп/с формируется медленными волнами, которые имеют значительную амплитуду при низких интенсивностях светового раздражения, при которых вызванные потенциалы типа первичных ответов практически незаметны. Поэтому, применив ритмическое световое раздражение с частотой около 5 имп/с, слабой интенсивности, можно наблюдать РП, образующуюся только из медленных волн, РП, состоящая из медленных волн РРП, может наблюдаться и за пределами зрительной коры (за пределами зоны регистрации первичных ответов на световые вспышки), но амплитуда ее здесь меньше, чем в проекционной зрительной области.
При рассмотрении РП не только на частоты, близкие к собственной частоте спонтанного α-подобного ритма и РРП, но и на более низкие и высокие частоты (2—10 Гц) можно видеть, что и в этих случаях основным элементом реакции являются медленные волны, идентичные волнам РРП.
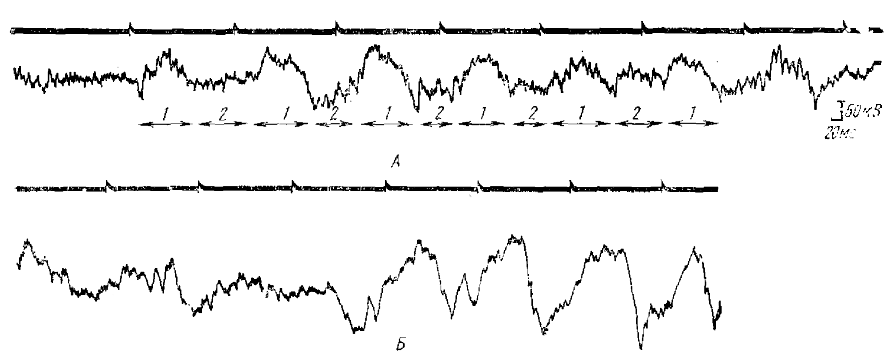
Если интервал между ритмическими световыми стимулами больше, чем длительность одной волны РРП, то такие волны также возникают после каждого стимула; причем длительность каждой волны почти не меняется. Поэтому кривая электрических потенциалов коры имеет характерный вид, состоит из медленных волн, электроотрицательных при регистрации от поверхности коры, которые разделены паузами (почти прямолинейными участками кривой), длительность которых равна разности между периодом следования световых стимулов и длительностью волн РРП (рис. 2, А). Иногда в этих «паузах» намечается появление второй волны РРП, но она имеет значительно меньшую амплитуду, чем первая волна, которая формирует основной рисунок реакции перестройки, как и при раздражении с частотой около 5 имп/с.
При более частых световых раздражениях, когда интервалы между стимулами становятся меньше, чем длительность одной волны РРП, форма кривой РП в затылочной коре кролика более сложна. Каждый световой стимул из ритмической серии в этом случае подается до окончания медленной волны, вызванной предыдущим стимулом. В результате этого наблюдается то же явление, которое было описано для РРП и спонтанных α-подобных волн. Если интервалы между стимулами таковы, что каждый стимул приходится на поверхностно-негативную вершину медленной волны, вызванной предыдущим стимулом, или на переход от негативной вершины к позитивной, то световые стимулы вызывают, помимо медленных волн, быстрые ответы высокой амплитуды — усиливающиеся потенциалы, часто во много раз превышающие по величине первичные ответы на одиночные световые раздражения той же интенсивности.
Далее, форма медленных волн, образующих РП, также усложняется при частоте световых стимулов, превышающей частоту волн РРП и спонтанных «-подобных волн. При одиночном световом раздражении (или раздражении зрительного нерва) первая волна РРП обычно начинается непосредственно медленным поверхностно-негативным отклонением, имеющим длительность 160—180 мс, которое, собственно, и образует волну. Лишь иногда между первичным ответом и медленным негативным отклонением наблюдается небольшая позитивная фаза длительностью до 50—60 мс (обозначим эти два компонента α-волны как следовую поверхностную позитивность и следовую поверхностную негативность). Характерным свойством следовой позитивности является ее способность к значительному увеличению амплитуды в том случае, если она вызывается раздражением, даваемым на фоне конца следовой негативности, вызванной предыдущим (обусловливающим) стимулом, т. е. при тех же условиях, при которых наблюдается возрастание амплитуды усиливающегося вызванного потенциала. Такое явление можно одинаково наблюдать при действии как парных, так и ритмических стимулов с интервалами порядка 120—- 160 мс. При парных раздражениях ответ на тестирующий стимул содержит высокоамплитудный (преимущественно поверхностно-позитивный), относительно быстрый усиливающийся вызванный потенциал; за ним следует также высокоамплитудное позитивное, но более медленное отклонение — следовая позитивность, которая непосредственно переходит в еще более медленную следовую негативность.
➥ Более подробно: Ритмические разряды последействия и α-подобная активность мозга животных
Следовая позитивность и следовая негативность формирует первую волну РРП, провоцируемого тестирующим стимулом. За ней может следовать еще несколько волн, но часто РРП оказывается редуцированным до одной первой волны. Такая же позитивно-негативная последовательность вызванных потенциалов возникает в ответ на каждый стимул и при ритмическом световом раздражении с частотой от 6 до 10 имп/с (кроме поздних волн РРП). РП, формирующаяся из таких потенциалов (рис. 2, Б), состоит из медленных позитивно-негативных комплексов (следовая позитивность и следовая негативность), чередующихся иногда с более быстрыми высокоамплитудными ответами — усиливающимися вызванными потенциалами (УО).
При частотах световых мельканий выше 10—12 имп/с РП описанного типа в затылочной коре кролика не возникает или возникает на очень короткое время (в самом начале ритмической стимуляции) и после нескольких вспышек света прекращается. Отсутствие РП при таких частотах становится понятным при рассмотрении вызванных потенциалов на парные стимулы с интервалом менее 80—100 мс. При интервалах, когда тестирующий стимул приходится на начало следовой негативности, вызванной обусловливающим раздражением, тестирующий стимул вообще почти не вызывает биоэлектрической реакции коры. За ним следует небольшое, часто едва заметное колебание потенциала (как показывают опыты с раздражением зрительного нерва и анализом распределения потенциалов по поперечнику коры, это колебание может быть отражением первичных синаптических процессов в глубоких слоях коры, Супин, 1966), но не возникает ни классического позитивно-негативного первичного ответа, ни усиливающегося ответа, ни следовой поверхностной позитивности или поверхностной негативности. Следовая негативность обусловливающей реакции развивается точно так же и имеет ту же форму, как и в отсутствие тестирующего стимула. Очевидно, следовая поверхностная негативность или, по крайней мере, ее начало сопровождаются процессом (или являются отражением процесса), который блокирует большинство корковых реакций на световые стимулы, даваемые на его фоне. Это, естественно, делает невозможным возникновение реакции перестройки описанного типа с частотой более 10—12 Гц.
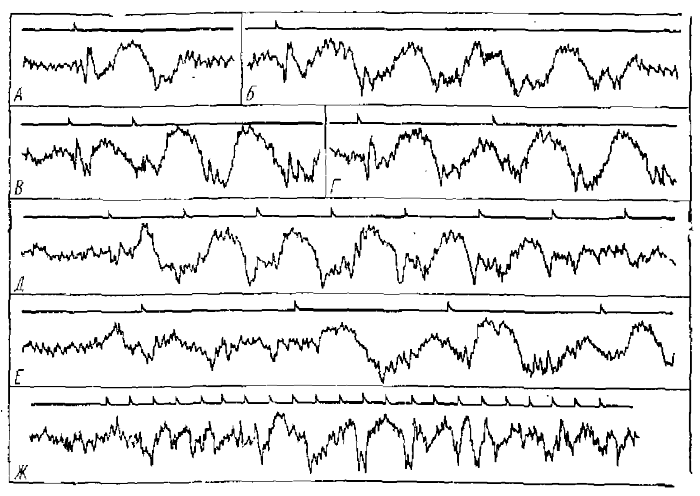
При частоте ритмической световой стимуляции 10 – 12 Гц медленные волны РРП все же могут формировать реакцию перестройки, но она имеет в этом случае частоту, не равную частоте стимуляции, а в два или (редко) в три раза меньшую. Принцип формирования реакции такого типа следующий: второй стимул ритмической серии раздражений не вызывает ответной реакции, так как попадает на начало следовой негативности, вызванной первым стимулом; третий стимул, попадая на конец этой негативности, вызывает реакцию, содержащую все компоненты, в том числе и новую негативную волну; поэтому четвертый стимул, попадающий на начало второй негативной волны, снова не вызывает реакции, а вызывает только ее пятый стимул, и т. д. (Супин, 1964). Таким путем возникает реакция, в которой медленные волны вызываются каждым вторым стимулом, т. е. наблюдается редукция частоты ритма (рис. 3). Аналогично может возникнуть при более высокой частоте раздражения реакция, содержащая одну волну на каждые три стимула, хотя такие случаи наблюдаются довольно редко (чаще при таких частотах раздражения стойкой реакции вообще не возникает).
При низких частотах ритмического светового раздражения (2—3 имп/с) может возникнуть реакция, в которой после каждого стимула успевает пройти не одна волна, а две (очень редко больше) волны РРП, т. е. частота реакции вдвое превышает частоту стимуляции (рис. 3).
Итак, медленные волны РРП играют основную роль в формировании РП ритма в коре мозга кролика при частотах, не слишком сильно отличающихся от собственной частоты РРП. Но, как было показано, РРП затылочной коры кролика можно считать искусственно спровоцированной вспышкой α-подобной активности, которая по своей природе должна мало отличаться от спонтанной α-подобной активности. Отсюда следует, что и медленные волны РП имеют ту же самую природу и являются волнами α-подобной активности, которая провоцируется и несколько видоизменяется ритмическими раздражениям с частотой, более или менее близкой к собственной частоте α-подобной активности.
Наблюдая за изменениями РРП по мере изменения частоты ритмических стимулов, сделано заключение об образовании РП из волн РРП; тем не менее это еще не может служить доказательством того, что природа регистрируемых медленных волн не изменяется при переходе от одиночных раздражений к ритмическим. Однако можно привести некоторые доказательства, подтверждающие α-подобную природу волн РП.
Прежде всего обращает на себя внимание четко выраженный частотный оптимум реакции, совпадающий с собственной частотой спонтанной и вызванной (РРП) α-подобной активности. РП описанного типа не может иметь частоту, сильно отличающуюся от частоты α-подобной активности (около 5 Гц), и если частота стимуляции значительно расходится с частотой α-подобной активности, то РП либо не возникает вовсе, либо возникает при явлении редукции или умножения частоты ритма, так что в конечном счете частота реакции оказывается все- таки довольно близкой к собственному α-подобному ритму. На сходство природы РП и α-подобной активности указывает также и одинаковое распространение по коре мозга (максимум в затылочной области и уменьшение амплитуды в соседних областях).
Наконец, для проверки идентичности механизмов волн α-подобной активности и волн РП можно применить тот же критерий, который использовался нами для установления аналогичности между спонтанной α-подобной активностью и РРП затылочной коры кролика, — критерий взаимодействия медленных волн с более быстрыми вызванными потенциалами коры. Такую проверку можно производить двумя способами. Во-первых, можно рассматривать взаимодействие медленных волн РП, вызываемой сильными ритмическими световыми стимулами, с быстрыми вызванными потенциалами, возникающими в ответ на эти же стимулы; различное фазовое соотношение быстрых вызванных потенциалов с медленными волнами можно получить меняя частоту ритмической стимуляции. Во-вторых, можно на фоне РП, вызванной относительно слабыми стимулами, давать дополнительные световые раздражения или раздражения зрительного нерва и рассматривать изменения ответов на эти внеочередные раздражения в зависимости от их фазового соотношения с волнами реакции перестройки.
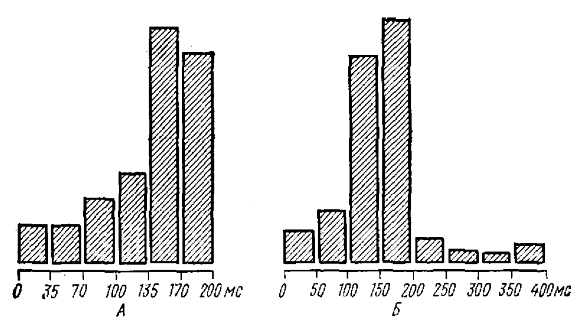
Оба способа дают вполне согласующиеся результаты (см. Гусельников, Супин, 1962). Стимул, совпадающий с концом поверхностно-негативной волны РП, вызывает высокоамплитудный ответ — усиливающийся вызванный потенциал. Стимул, совпадающий с позитивной вершиной или началом поверхностно-негативной волны РП вообще не вызывает ответа или вызывает ответ минимальной амплитуды. Характер изменения амплитуды вызванных потенциалов в зависимости от фазового соотношения с медленной волной полностью совпадает как для волн спонтанной α-подобной активности и α-подобного РРП, так и для волн РП (рис. 4). Это служит еще одним подтверждением взгляда, согласно которому рассмотренный здесь тип РП является искусственно вызываемой α-подобной активностью.

Значительным подкреплением наших данных явились результаты исследований В. Б. Полянского (1966) по изучению РП в коре кролика на нейронном уровне. Он показал, что каждое световое раздражение из ритмической серий вызывает в нейронах коры (рис. 5) короткую первоначальную вспышку активности, за которой следует длительная тормозная пауза. Этой паузе в суммарной электрической активности соответствует медленная поверхностно-негативная (глубинно-позитивная) волна. Если интервалы между ритмическими стимулами больше, чем длительность медленной волны, то в конце каждой волны можно наблюдать возобновление импульсной активности нейронов, наступающей еще до вспышки активности, вызываемой следующим стимулом. Если интервалы меньше, чем длительность медленной волны и тормозной паузы, то каждый стимул может вызвать вспышку импульсной активности, и тем меньшую, чем меньше интервал между стимулами. При относительно высокой частоте ритмической афферентной стимуляции, когда каждый стимул приходится на фазу глубокого торможения, вызванного предыдущим стимулом, РП не возникает, включение ритмической стимуляции затормаживает нейроны.
Эти данные вполне коррелируют с нашими данными, приведенными в связи с анализом генезиса α-волны зрительной коры кролика и ее тормозной природы. Значит, действительно РП на низкую частоту раздражения является искусственно вызываемой α-подобной активностью.

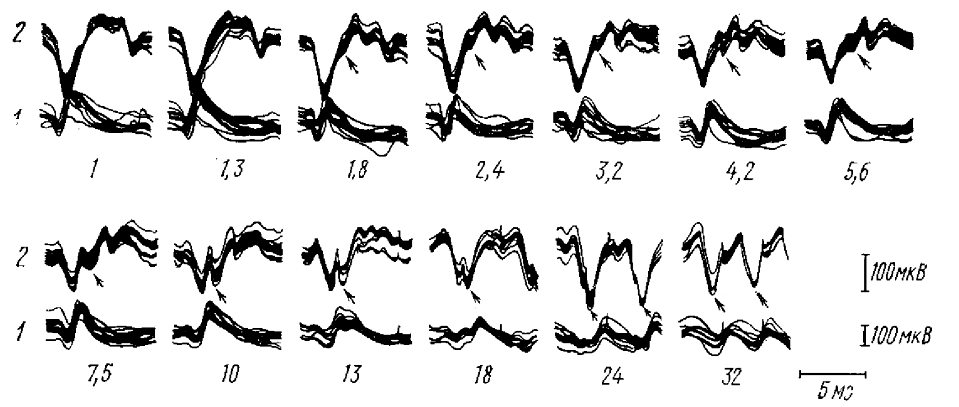
В какой степени этот вывод, полученный при исследовании РП и ЭЭГ кролика, может быть справедлив и для РП в ЭГГ человека? Пока нет точных доказательств аналогичности РП кролика и человека. Но детальный анализ формы осцилограмм дает основание предполагать, что такая аналогичность вероятна. Несмотря на вариабельность формы потенциалов РП, регистрируемых у разных испытуемых, можно выделить несколько основных свойств, показывающих, что формирование каждой волны РП у человека (рис. 6) происходит из тех же компонентов, что и формирование волн РП в затылочной коре кролика. Вызванные потенциалы сложной формы, возникающие в ЭЭГ человека при действии редких световых стимулов, сменяются при увеличении частоты стимуляции более сглаженными негативными волнами, длительность которых близка к длительности волн α-ритма; амплитуда этих волн увеличивается по мере приближения частоты стимуляции к частоте α-ритма (см. рис. 6). Когда частота стимуляции превышает частоту α-ритма, негативные волны сменяются типичными позитивно-негативными комплексами; при дальнейшем увеличении частоты стимуляции амплитуда волн РП резко падает, иногда до полного ее отсутствия (Ciganek, 1958, 1961; Werre, Smith, 1964). Форма медленных волн РП у человека может значительно-искажаться более быстрыми вызванными потенциалами. Видимо, можно полагать, что РП и в ЭЭГ человека формируется на основе вызванных α-волн, т. е. тех же механизмов, что и в ЭЭГ кролика. Эти данные совпадают с мнением М. Н. Ливанова (1944), что РП есть результат усиления той частоты из числа существующих спонтанных, которая наиболее близка к частоте ритмической световой стимуляции.
Итак, мы выяснили, что РП, возникающая на основе вызванных α-волн или α-подобных волн, не может существовать без редукции ритма при частотах стимуляции, превышающих в 1,5— 2 раза собственную частоту α-активности (α-подобной активности).
Реакция перестройки ритма на высокие частоты стимуляции. Известно, что РП на ритмическое световое раздражение может наблюдаться и при значительно более высоких частотах стимуляции. Очевидно, в этих случаях РП имеет иной механизм, скорее всего является последовательностью вызванных потенциалов, устойчивых к частой повторной стимуляции. На различные механизмы РП в диапазоне α-активности и в более высокочастотном диапазоне указывает даже форма частотно-амплитудных характеристик РП. И у человека, и у ненаркотизированных животных зависимость амплитуды волн РП на ритмический свет от частоты стимуляции имеет в принципе одинаковый характер. Одна вершина (максимальная амплитуда волн реакции) наблюдается на частоте, равной собственной частоте α-активности (α-подобной активности) или слегка превышающей ее, она соответствует реакции перестройки α-подобного типа. Вторая вершина наблюдается при значительно более высоких частотах — порядка 20 Гц или выше (см. рис. 8). В промежутке между двумя оптимумами лежит полоса меньшей амплитуды РП. Это явление может быть выражено настолько четко, что в определенном диапазоне частот ритмические световые раздражения вообще не вызывают заметной реакции, хотя при более низких и более высоких частотах РП имеет значительную амплитуду. Поскольку при парных раздражениях не обнаруживалось двух диапазонов интервалов между стимулами, при которых тестирующий стимул вызывал бы α-волну значительной амплитуды, а, наоборот, показано, что при коротких интервалах тестирующий стимул вообще не вызывает α-волны, можно заключить, что возобновление РП на ритмический свет при повышении частоты стимуляции связано не с вызванными α-подобными волнами, а с ответами другого типа. Изучение циклов восстановления вызванных потенциалов различного типа (см. Гусельников, Супин, 1968) показывает, что при таких частотах стимуляции могут возникать только ПО. Однако более тщательные исследования показывают, что, хотя это и верно, ПО в таких условиях очень сильно отличаются от ПО, регистрируемых в тех же условиях на одиночные световые раздражения.
Сопоставим реакции зрительной коры и проводящих путей зрительного анализатора (Супин, 1966) (зрительный нерв или зрительный перекрест) на ритмические световые вспышки различной частоты (рис. 7). При редких световых вспышках электрический ответ зрительного перекреста на каждую вспышку состоит из двухвершинного позитивного потенциала (ответа на включение) и небольшого потенциала, расцениваемого как ответ на выключение (Bartley, Bishop, 1942; Bartley, 1942). В ответе на включение при редких вспышках основным является первый пик, второй пик имеет меньшую амплитуду или полностью маскируется первым. Эти два пика должны отражать прохождение по зрительному нерву двух залпов импульсов, генерируемых сетчаткой в ответ на каждую вспышку света; причем первый залп является значительно более мощным. Соответственно этому ПО коры на одиночную световую вспышку имеет вид не простого позитивно-негативного колебания, а более сложную форму — фактически он является наложением двух ПО, вызываемых двумя залпами импульсов из сетчатки; причем основным является первый ответ, а второй создает небольшое дополнительное колебание на его негативной фазе. Третий пик — эффект выключения — при коротких световых вспышках, обычно применяющихся при исследовании вызванных потенциалов, имеет очень небольшую величину, и в коре не наблюдается ответа заметной величины, соответствующего этому пику. Поэтому мы будем рассматривать только изменения двух компонентов эффекта включения и соответствующих им корковых ПО. Для более четкой терминологии мы будем обозначать компонент коркового ПО, вызываемого первым залпом импульсов из сетчатки, как компонент I, а ответ на второй залп — как компонент II (см. рис. 7).
При учащении световых вспышек наблюдается следующее характерное явление. Первый пик ответа зрительного перекреста по мере учащения раздражений сильно уменьшается, а второй пик увеличивается, в его латентный период укорачивается. Поэтому при частотах 20—30 имп/с и более почти весь ответ зрительного перекреста образован вторым пиком, а редуцированный первый пик наблюдается лишь в виде небольшого перегиба на переднем фронте второго. Аналогичные изменения происходят и в коре мозга: ответ на второй залп импульсов из сетчатки (компонент II), который при редких стимулах выявлялся только как небольшое колебание на негативной фазе первого ответа, при учащении вспышек начинает вырастать параллельно с ростом второго пика ответа зрительного перекреста, а компонент I ответа коры уменьшается вместе с уменьшением амплитуды первого пика ответа зрительного перекреста, так что при частотах раздражения около 10—15 имп/с величины обоих компонентов коркового ответа выравниваются, а при более высоких частотах практически весь ответ коры образован компонентом II, а компонент I почти не обнаруживается. Таким образом, выявляется первое основное различие между ПО зрительной коры на редкие и на частые световые вспышки: при редких вспышках регистрируется почти исключительно ответ на первый залп импульсов, посылаемых сетчаткой, а при частых — ответ на второй залп импульсов.
При сопоставлении временных соотношений между ответами зрительного перекреста и коры на редкие и частые световые вспышки видно, что реакция коры на частые световые вспышки должна состоять преимущественно из последовательности негативных фаз ПО на второй залп импульсов из сетчатки. Это кажется довольно неожиданным, поскольку поверхностно-негативная фаза ПО имеет значительно меньшую скорость восстановления после предыдущего стимула, чем поверхностно-позитивная. Кроме того, вывод, сделанный только на основании прослеживания перехода от одиночных вызванных потенциалов к ритмическим, может вызвать вполне обоснованные сомнения. Поэтому было проведено (Супин, 1964) специальное исследование вопроса об участии поверхностно-позитивной и поверхностно-негативной фаз ПО в формировании РП на частые вспышки света. Изучалось послойное распределение по поперечнику коры и чувствительность к различным воздействиям реакции на частые световые вспышки в сравнении с теми же свойствами фаз ПО на одиночные вспышки.
Известно, что при погружении регистрирующего электрода от поверхности в глубину коры наблюдается постепенное уменьшение латентного периода негативной фазы ПО, что свидетельствует о движении возбуждения от глубоких слоев коры, к которым приходят специфические афференты, к поверхностным слоям; это вертикальное движение возбуждения и заканчивается активацией (деполяризацией) поверхностных элементов коры, находящей отражение в поверхностно-негативной фазе ПО. Для поверхностно-позитивной фазы ПО такое вертикальное движение не описано. Это различие было использовано для установления преимущественного сходства реакции на частые световые вспышки с поверхностно-позитивной или поверхностно-негативной фазой ПО. Если реакция состоит из потенциалов, аналогичных поверхностно-позитивной фазе ПО, то при погружении регистрирующего электрода в глубину коры не должны изменяться латентные периоды ее колебаний, а если реакция родственна поверхностно-негативной фазе ПО, то изменение латентных периодов ее колебаний должно наблюдаться. Естественно, какой бы точки зрения ни придерживаться относительно генеза отрицательной и положительной фаз ПО коры, это предположение пока правомочно.
➥ Более подробно: Вторичные ответы коры больших полушарий
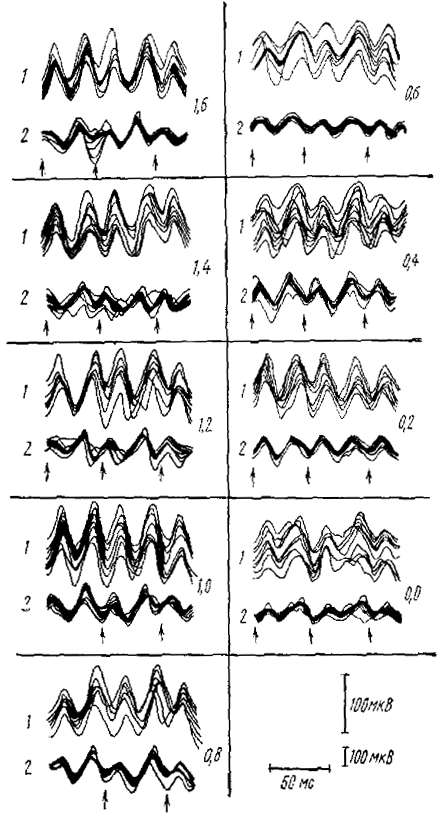
Опыты А. Я. Супина показали, что при одновременной регистрации реакции на частые световые мелькания от поверхности и от разных слоев коры мозга кошки, по мере погружения глубинного регистрирующего электрода в кору отводимая им реакция все больше смещается во времени относительно реакции, регистрируемой от поверхности, и опережает ее (см. рис. 8). Это опережение в глубоких слоях коры (1,6—1,8 мм) может достигать 7—8 мс. Следовательно, реакция коры на частые световые вспышки отражает не только локальные процессы на поверхности или в глубине коры, а процессы, которым свойственно движение из глубоких слоев коры к поверхности, т. е. является более близким аналогом поверхностно-негативной фазы ПО.
Этот вывод подтверждается также и опытами с влиянием на исследуемую реакцию асфиксии и аппликации на поверхность коры новокаина. Опыты ставились на основе данных, согласно которым поверхностно-позитивная и поверхностно-негативная фазы ПО имеют разную чувствительность к действию фармакологических веществ и к аноксии. Мы не будем пытаться связывать такую различную чувствительность с природой фаз ПО. Для рассмотрения интересующего нас вопроса достаточно факта, что это различие существует. Сравним по этому признаку реакцию на частые световые мелькания с фазами ПО.

ненаркотизированной кошки, возникающих на световые вспышки, при аппликации на поверхность коры 2%-ного раствора новокаина. А — ответы на световые вспышки с частотой 2 имп/с; Б — с частотой 20 имп/с (по Супину, 1966)
При аппликации на поверхность коры раствора новокаина ПО претерпевают отчетливые изменения. Особенно интересна начальная стадия (неглубокая новокаинизация коры), когда наблюдается заметное уменьшение поверхностно-негативной фазы ответа при одновременном увеличении поверхностно-позитивной фазы (см. рис. 9). Реакция на частые световые вспышки в это время заметно уменьшается по амплитуде. Более того, параллельно с ростом поверхностно-позитивной фазы ПО на одиночные вспышки, в реакции на частые вспышки выявляется новый компонент—короткие поверхностно-позитивные колебания, разделенные паузами. Очевидно, эти результаты нельзя интерпретировать иначе, как приняв, что в норме реакция на частые световые вспышки образована последовательностью вызванных потенциалов, сходных с поверхностно-негативной фазой ПО, а поверхностно-позитивные фазы в образовании этой реакции заметного участия не принимают и выявляются только при новокаинизации коры.
Аналогичные результаты были получены и в опытах с непродолжительной асфиксией путем выключения искусственного дыхания у кураризированных кошек. В опытах с применением асфиксии наблюдается явление, дающее дополнительные сведения о природе реакции зрительной коры на частые световые мелькания. Два компонента ПО, вызываемые двумя залпами импульсов из сетчатки (компоненты I и II, см. рис. 8), оказываются различно чувствительными к асфиксии. Ответ на второй залп импульсов (компонент II) исчезает значительно раньше, чем ответ на первый залп (компонент I). так что на определенной стадии асфиксии (рис. 10, А— 60 с) в коре наблюдается ПО просто двухфазной формы, латентный период которого указывает на то, что он вызван первым залпом импульсов, т. е. является компонентом I. Именно в это время (рис. 10, В — 70 с) возникает блокада реакции коры на частые световые стимулы, что является дополнительным подтверждением того, что эта реакция состоят из компонентов II кортикальных ответов, вызываемых каждой световой вспышкой.
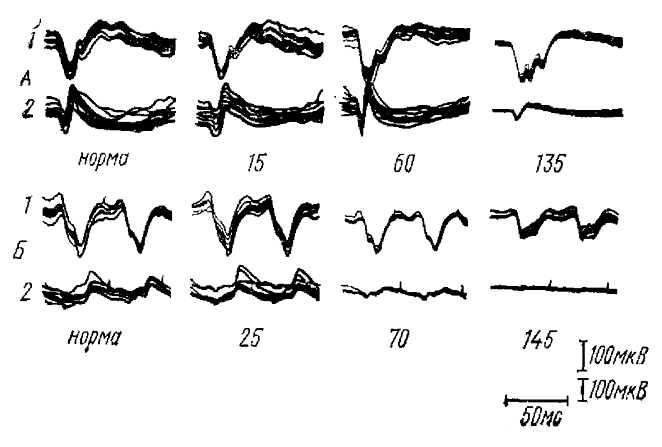
Разновременная блокада при асфиксии двух компонентов ПО дает сведения и о корковой природе реакции на частые световые стимулы. Блокада компонента II кортикального ответа наблюдается на такой стадии асфиксии, когда ответы зрительного перекреста еще практически не изменены, т. е. не изменена форма залпов импульсов, посылаемых сетчаткой. Таким образом, блокада при асфиксии компонента II ПО происходит не за счет периферических процессов, а в результате процессов в центральных отделах анализатора, вероятнее всего, в самой коре. Таким образом, два компонента ПО не являются совершенно однотипными реакциями на два последовательных афферентных залпа, — свойства их в значительной степени различны. Возможно, разные афферентные залпы импульсов адресуются к разным элементам коры.
На примере рассмотренной реакции хорошо видно сильное различие между ответами на частые ритмические стимулы и на одиночные или парные стимулы. Конечно, и при переходе от одиночных стимулов к парным можно видеть изменения, во многом сходные с изменениями при переходе от одиночных стимулов к ритмическим. Так, при укорочении интервалов между парными световыми стимулами хорошо видно замещение первого компонента ответа на тестирующий стимул вторым компонентом, хотя характер ответов в этом случае не такой, как при ритмических раздражениях, — основой является не поверхностно-негативная фаза ПО, а поверхностно-позитивная фаза ПО или усиливающегося ответа. Но даже этот процесс выражен значительно сильнее при ритмических раздражениях, а преобладание поверхностно-негативной фазы над поверхностно-позитивной при малых интервалах между стимулами вообще не наблюдается при парных раздражениях и свойственно только реакции на ритмическую стимуляцию. Очевидно, в процессе ритмической стимуляции происходит изменение цикла восстановления поверхностно-негативной фазы ПО, в результате чего скорость се восстановления становится не только ниже, но значительно выше, чем у поверхностно-позитивной фазы. Можно предположить, что это происходит под влиянием неспецифической ретикулярной активации, вызываемой тоническим активирующим воздействием длительной серии частых световых вспышек. В пользу такого предположения свидетельствует то, что амплитуда ответов на частые (8—11 в 1 с. и выше) ритмические световые вспышки увеличивается при электрическом раздражении ретикулярной формации среднего мозга (причем этот эффект блокируется нембуталовым наркозом) и уменьшается при введении животному аминазина, тогда как ПО, вызванные редкими вспышками, при этом почти не изменяются (Супин, 1963).
Что касается участия в РП вторичных ответов коры, то имеются только данные относительно позднего вызванного потенциала зрительной коры кролика. На возможность участия этого вызванного потенциала в образовании РП на ритмические световые раздражения указывали Р. М. Мещерский и Г. Д. Смирнов (1961), Р. М. Мещерский и др. (1962). Однако надо полагать, что, поскольку поздние вызванные потенциалы зрительной коры кролика тесно связаны с появлением α-подобных волн, последовательность этих вызванных потенциалов следует скорее считать на самостоятельной ритмической реакцией, а компонентом РП ритма α-подобного типа (См. Гусельников, Супин, 1968).
Ю. Ф. Педанов (1971) показал, что РП на низкочастотное раздражение более генерализована по коре, чем РП на высокие частоты раздражения. В последнем случае она приурочена к проекционной зоне коры и зависит от изолированного вовлечения в процесс навязывания только зрительных релейных ядер, что совпадает с данными о роли специфических путей в высокочастотной РП, приведенными выше.
По данным (Гусельников, Супин, 1968), обе реакции перестройки по-разному зависят от одних и тех же сдвигов функционального состояния мозга. Низкочастотная РП лучше выражена при среднем уровне активации коры и подавляется при возбуждении и при сне, высокочастотная РП имеет максимальную величину при наибольшем уровне активации.
Отсюда, видимо, и большой разнобой и противоречия в сведениях относительно вопроса о зависимости РП от функционального состояния коры. Именно в связи с этим возникает вопрос, насколько понятие лабильности приложимо в данном случае к такому сложному образованию, как кора.
Исследователи легко связывают изменение реакции перестройки с уровнем лабильности коры, вообще, не акцентируя внимания на том, с лабильностью каких элементов, звеньев нервной сети коры имеет дело экспериментатор (учитывая, что понятие лабильности было четко определено Н. Е. Введенским).
Видимо, в каждом конкретном случае, когда исследователь пытается использовать РП как определенный показатель тех или иных функций или функциональных особенностей мозга, и прежде всего коры больших полушарий, совершенно необходимо точно знать, с какими конкретными реакциями он имеет дело и каков тонкий генез этих реакций.
В связи с этим гармонический анализ РП в ЭЭГ с использованием любых простых или сложных электронных анализаторов или математические приемы исследования были бы более правомерными, если бы мы точно знали, с какими нервными процессами и с какими нейронными структурами мозга мы имеем дело.
Отсюда следует, что еще остается крайне желательным дальнейшее, более глубокое исследование генеза этой интересной реакции мозга — реакции, которая при более правильном подходе даст значительно больше для вскрытия важных механизмов мозга животных и человека, чем тот эмпирический подход в использовании РП, который имел и еще иногда имеет место.