Целый ряд ЭЭГ феноменов, возникающих на те или иные виды раздражений, являются не чем иным, как отражением активности механизмов, которые в норме генерируют спонтанную активность мозга. Это касается прежде всего ритмического разряда последействия (РРП), реакции усиления (РУ) и реакции вовлечения (РВ).
У бодрствующего человека РРП в ЭЭГ совпадает по частоте, локализации и форме волн со спонтанным ритмом коры. Считается, что РРП у человека является искусственно вызванной вспышкой α-ритма (Gastaut, Hunter, 1950, и др.). Это подтверждается и другими данными. Так, у испытуемых с большей частотой α-ритма больше и частота РРП. При засыпании, в стадии, когда α-ритм исчезает, нельзя вызвать и РРП, он возникает также только у тех испытуемых, у которых хорошо выражен и α-ритм (Peacock, 1970).
У животных РРП, которые приурочены к новой коре и таламусу, также, видимо, являются большей частью аналогами одного из двух основных ритмов, в генерации которых кора и таламус играют решающую роль — α-подобной и веретенообразной активности.
Исследования, связанные с изучением генеза РРП, были проведены в нашей лаборатории (Гусельников, Супин, 1962, 1968; Супин, 1964). Подвергся изучению РРП затылочной коры бодрствующего кролика, вызываемый афферентным зрительным раздражением. В зрительной коре мозга кролика помимо ПО при афферентном раздражении (или «дендритного» — при прямом раздражении коры током) возникает медленная поверхностно-негативная волна длительностью 150—200 мс, за которой следует еще один вызванный потенциал, описываемый разными исследователями как «экстра-потенциал» (Мещерский, Смирнов, 1961), или вторичный ответ зрительной коры кролика (Иваницкий, 1966; Fuster, Docter, 1962).
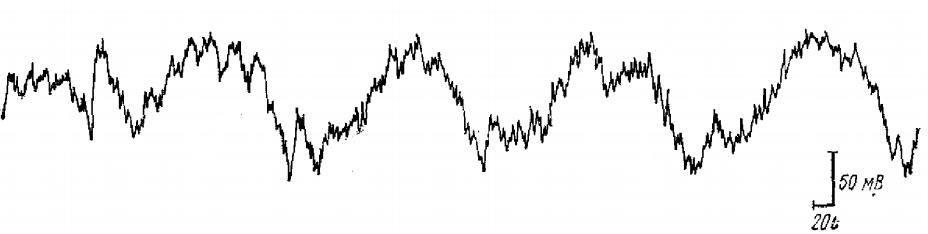
НВ описывается часто как компонент реакции на афферентное раздражение. Однако, как правило, при хорошем функциональном состоянии животного и особенно в условиях хронического эксперимента возникает несколько таких медленных поверхностно-негативных волн от 2 до 5 и более, которые и составляют РРП зрительной коры кролика (рис. 1). «Экстра-потенциалы» возникают после каждой волны такого разряда. Таким образом, НВ следующая за первичным ответом коры, является лишь первой волной РРП.
РРП в зрительной коре кролика имеет ряд признаков, сближающих его со спонтанной ритмической активностью затылочной коры — α-подобным ритмом кролика. Он имеет частоту, близкую к частоте спонтанного α-подобного ритма (порядка 5 в 1 с), сходную форму волн. Локализация его в коре совпадает с локализацией спонтанной α-подобной активности. Несмотря на большое сходство РРП зрительной коры кролика со спонтанной α-подобной активностью, для строгого доказательства их аналогичности этого недостаточно. Поэтому было необходимо найти какой-то критерий сравнения РРП и α-подобной активности.
Нами в качестве такого критерия был выбран характер взаимодействия волн РРП и спонтанных α-подобных волн с вызванными потенциалами коры, возникающими на световое раздражение сетчатки или раздражение зрительного нерва (Гусельников, Супин, 1962).
При исследовании этого явления было обнаружено, что в этом случае зависимость вызванного потенциала зрительной коры кролика от фазы спонтанной α-подобной активности (или РРП) выражена чрезвычайно резко (рис. 2). Ответы максимальной величины (они возникают на фазе каждой α-подобной волны или волны РРП, соответствующей переходу от поверхностно-негативной вершины волны к электропозитивной) могут в десятки раз превышать ответы минимальной величины (на позитивной вершине волны и на переходе от позитивной вершины к негативной) (рис. 2, Г). Эта закономерность характерна для ответов на раздражение сетчатки короткой вспышкой света (Гусельников и Супин, 1962; Pearlman, 1963) и для ответов на электрическую стимуляцию зрительного нерва или латерального коленчатого тела, причем изменения ответа циклически повторяются, совпадая с волнами РРП.
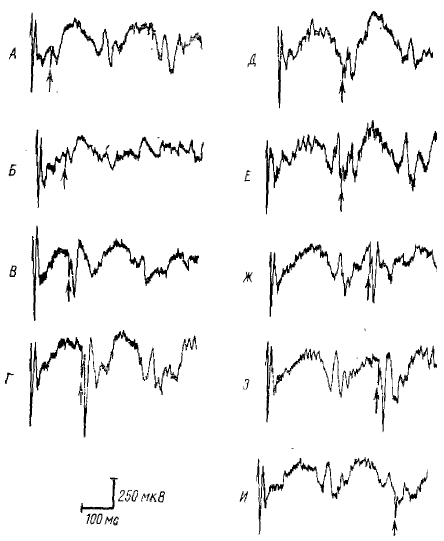
Детальное исследование показало, что способностью к резкому изменению амплитуды в зависимости от фазы фоновой α-подобной волны или волны РРП обладает не классический ПО, а другой вид вызванного потенциала. На обычном фоне, не содержащем α-подобных волн, этот вызванный потенциал имеет небольшую амплитуду. Но он резко усиливается, если совпадает с концом (переходом от негативной вершины к позитивной) каждой α-подобной волны, и почти полностью блокируется, если совпадает с ее началом (позитивной вершиной и переходом от позитивной вершины к негативной) (Гусельников, Супин, 1962). За такую особенность этот вид вызванного потенциала был назван «усиливающимся» ответом (УО).
ПО также изменяется на фоне РРП — вначале почти полностью (или полностью) блокируется, а затем постепенно восстанавливается. Но эти изменения, возможно, связаны не с РРП, а с каким-либо другим сопутствующим ему процессом, который запускается тем же обусловливающим стимулом, что и РРП. Об этом говорит ход восстановления ПО после обусловливающего стимула: восстановление имеет большую длительность (порядка секунды) и не обнаруживает заметной цикличности, связанной с волнами РРП. Наоборот, УО совершенно одинаковым образом изменяется на каждой волне РРП в зависимости от фазового соотношения с этими волнами (рис. 3).
УО имеет несколько большую длительность, чем ПО (см. рис. 2, 3), но это различие можно выявить только в том случае, если ответы вызываются электрическим раздражением зрительного нерва (или тракта) или латерального коленчатого тела, когда в кору поступает компактный, не растянутый во времени афферентный залп. Вследствие этого ответы имеют достаточно малую длительность. Общая длительность ПО зрительной коры кролика в этих условиях обычно не превышает 20 мс (при регистрации от поверхности коры позитивная фаза достигает вершины через 6—7 мс после раздражения зрительного нерва, негативная фаза через 9—11 мс), а длительность УО составляет около 30 мс (вершины поверхностно-позитивной и поверхностно-негативной фаз приходятся соответственно на 11 —14 и 20—23 мс). Уже эти расхождения временных характеристик указывают, что имеют место не две модификации одного и того же ответа, а два различных вызванных потенциала. Это же доказывается и сравнением формы вызванных потенциалов, возникающих на разных фазах волны РРП. При достаточно больших интервалах между обусловливающим стимулом, вызывающим РРП, и тестирующим стимулом, когда тестирующий стимул дается после первой волны РРП, ответ на тестирующий стимул снова приобретает характер ПО как на одиночное раздражение. Однако переход от УО к ПО при постоянном отставлении тестирующего стимула происходит путем уменьшения УО и одновременного постепенного нарастания ПО, так что при определенных интервалах между обусловливающим и тестирующим стимулом (на рис. 3—220 мс) ответ на тестирующий стимул имеет четыре вершины: две поверхностно-позитивных, соответствующих по времени позитивным вершинам первичного и УО, и две негативных, соответствующих негативным вершинам ПО и УО.

Итак, фоновые α-подобные волны зрительной коры кролика и волны РРП обладают свойством резко изменять амплитуду УО в зависимости от фазы волны, на которую приходится ответ. С целью использования этого свойства в качестве характерного признака α-подобной активности, пригодного для ее идентификации, необходимо было показать, что это свойство является исключительной особенностью α-подобной активности и ее модификацией и не характерно для остальных ритмов.
Эксперименты показали, что только α-подобному фоновому ритму свойственна способность резко изменять амплитуду вызванных потенциалов, возникающих на разных фазах его волн. Другие исследованные формы ритмической фоновой активности — веретенообразные колебания, стресс-ритм, а также нерегулярная фоновая активность — тоже влияли на величину вызванных потенциалов, но это влияние было слабее и обнаруживалось только путем статистического анализа большого количества измерений амплитуд ответов, возникающих на разных фазах фоновых потенциалов.
Но это же свойство (резкое изменение амплитуды УО в зависимости от фазового соотношения с волнами, на фоне которых он возникает), наблюдаемое только для α-подобной активности, наблюдается также и для РРП затылочной коры кролика. Отсюда можно предположить, что эти два вида активности аналогичны по своей природе, т. е. РРП является искусственно спровоцированной вспышкой α-подобной активности.
Связь спонтанной и вызванной α-подобной активности (т. е. РРП) с процессами в одиночных элементах коры
Рассмотрев вызванную активность — РРП затылочной коры кролика — и установив идентичность его механизмов механизмам спонтанного α-подобного ритма кролика, попытаемся на примере РРП выяснить нейрональные механизмы спонтанной α-подобной активности, так как только для этого аналога α-ритма имеются достаточно подробные экспериментальные данные. Возможность искусственного вызова РРП делает его для этой цели намного более удобным объектом исследования, чем сама спонтанная α-подобная активность.
Рассмотрим сначала процессы в одиночных нейронах коры во время РРП, вызванного одиночным световым раздражением сетчатки или электрическим раздражением зрительного нерва.
В дальнейшем мы будем опираться на данные, полученные в условиях полностью выраженного РРП, и результаты работ, в которых возникала только одиночная α-подобная волна из всего РРП (медленная негативная волна, НВ). Во всех случаях свойства этой первой одиночной и последующих α-волн оказывались одинаковыми, а максимальная стабильность именно первой волны представляет наибольшие удобства для исследования.
Вызванная α-подобная волна затылочной коры кролика состоит из двух основных компонентов, которые при регистрации от поверхности коры имеют вид более короткой поверхностно-позитивной волны (ПВ) и длительной (около 200 мс) поверхностно-негативной волны (НВ). В большинстве случаев поверхностно-позитивный компонент α-подобной волны, вызванной афферентным раздражением, бывает плохо выражен и нередко полностью маскируется НВ, начинающейся вслед за ПО.
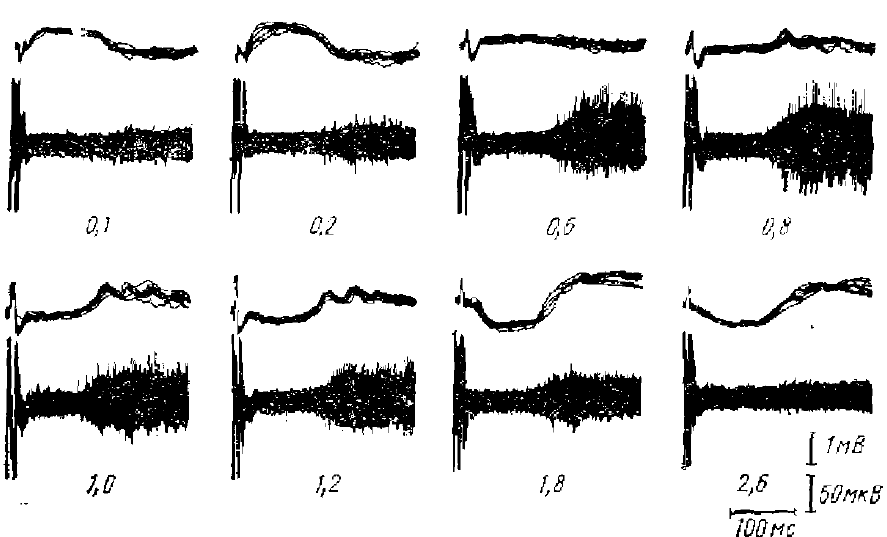
Сопоставление импульсной активности одиночных нейронов коры с вызванной α-подобной волной показывает, что НВ сопровождается сильным угнетением нейронной активности (Кондратьева, 1964; Полянский, 1965; Супин, 1966; Гусельников, Супин, 1968). В большинстве случаев наблюдается полное отсутствие импульсных разрядов во время НВ. Хотя большинство нейронов до раздражения имеют значительную фоновую активность, а после окончания НВ (если за ней не следуют повторные волны РРП) — импульсные разряды нейронов снова возобновляются. Импульсная активность во время НВ не обнаруживается не только при регистрации активности одиночных нейронов, но и при регистрации «шумовой» импульсной активности. Это значит, что происходит подавление импульсной активности не отдельных корковых нейронов, а подавляющей массы нейронов, имеющих спонтанную активность; причем угнетается активность по всей толще коры — от поверхности до наиболее глубоких ее слоев (рис. 4).
Эти факты показывают, что НВ (т. е. α-подобная волна) затылочной коры кролика сопровождается тормозным процессом, охватывающим массу нейронов всех слоев коры. Очевидно, афферентное раздражение вызывает в корковых нейронах ТПСП, длительность и латентный период которых близки к длительности и к латентному периоду НВ.
Существование длительного ТПСП в корковых нейронах во время α-подобной волны подтверждается и исследованием импульсных реакций нейронов на повторные раздражения, даваемые на фоне α-подобной волны, вызванной предыдущим (обусловливающим) стимулом (Полянский, 1965; Супин, 1966). Во время α-подобной волны блокируется нe только фоновая активность нейронов, но и полностью отсутствуют (в начале волны) реакции нейронов на афферентные раздражения (рис. 5). Реакция нейронов возобновляется только тогда, когда повторный (тестирующий) стимул совпадает с концом негативной α-подобной волны, вызванной предыдущим (обусловливающим) стимулом. Причем, чем ближе к концу α-подобной волны дается тестирующее раздражение, тем более интенсивна импульсная реакция нейронов (рис. 5). Минимальный интервал, при котором тестирующее раздражение зрительного нерва вызывает ответную импульсную активность, — около 60—70 мс. При увеличении интервала между обусловливающим и тестирующим стимулами сверх этой величины наблюдается увеличение интенсивности ответной импульсной реакции нейронов на тестирующее раздражение. Максимальная активация нейронов коры тестирующим стимулом наблюдается в том случае, если он дается непосредственно перед тем моментом, когда должна кончиться пауза в импульсной активности нейрона, вызванная обусловливающим стимулом (интервал 120—140 мс), при таких условиях интенсивность реакции нейрона в несколько раз выше, чем при одиночном раздражении (см. рис. 5). Если интервал между стимулами более 120— 140 мс и тестирующий стимул дается на фоне импульсной активности нейрона, возобновившейся после тормозной паузы, то реакция на тестирующий стимул заметно уменьшается.
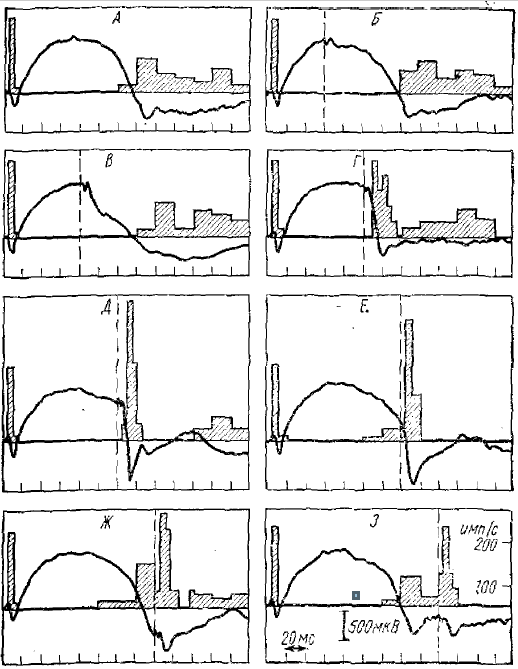
Эти факты согласуются с гипотезой, в которой обусловливающее афферентное раздражение должно вызвать длительный и высокоамплитудный ТПСП в нейронах коры. После обусловливающего стимула гиперполяризация нейронов еще сильна и разряды нейронов отсутствуют и не могут быть вызваны активацией деполяризующих синапсов через афферентные волокна. Через 60—70 мс гиперполяризация несколько спадает, так что при повторной афферентной активации появляется слабая вспышка импульсной активности. При еще больших интервалах между стимулами гиперполяризация еще сильнее уменьшается к моменту тестирующего раздражения, что вызывает и более интенсивную импульсную реакцию. Максимальная реакция должна наблюдаться в том случае, если тестирующее раздражение дается в момент, когда вызванный обусловливающим раздражением ТПСП закончится совсем, перед возобновлением или в самый момент возобновления импульсных разрядов после предшествующей паузы.
Именно это, как было показано выше, и наблюдается в действительности.
Вопрос о том, почему реакция нейронов в этот момент оказывается значительно интенсивнее, чем реакция на одиночное раздражение, возникающая до начала ТПСП, а также почему тестирующая реакция уменьшается на фоне возобновившейся после тормозной паузы импульсной активности, мы пока не рассматриваем. К этому вопросу мы вернемся при обсуждении возможной причины наблюдаемого постсинаптического торможения.
Показанная зависимость импульсных реакций нейронов на тестирующее раздражение от соотношения с α-подобной волной, вызванной обусловливающим раздражением, может в значительной степени объяснить взаимодействие α-подобной волны с вызванными потенциалами коры, которое, как было показано ранее, является характерным признаком α-подобной активности коры головного мозга кролика. Действительно, сопоставление циклов восстановления вызванных потенциалов и импульсных реакций коры показывает, что при изменении интервала между тестирующим и обусловливающим стимулами наблюдается совершенно четкий параллелизм изменений импульсных реакций на тестирующее раздражение и УО коры (рис. 6), У большинства нейронов минимальный интервал между стимулами, при котором возникает тестирующая импульсная реакция, очень близко совпадает с интервалом, при котором появляются первые признаки УО. Увеличение интервала между стимулами сопровождается параллельным увеличением интенсивности импульсной реакции и амплитуды УО, максимальная интенсивность импульсной реакции также наблюдается при таком же интервале между стимулами, при котором УО имеет максимальную амплитуду. Наконец, дальнейшее увеличение интервала ведет к параллельному ослаблению импульсной реакции нейронов и уменьшению амплитуды УО на тестирующее раздражение. Такая тесная связь между двумя процессами указывает на их вероятную причинную зависимость. Сопоставление формы и длительности УО с временным распределением импульсных разрядов нейрона на афферентное раздражение показывает, что УО является следствием импульсной реакции, а по наоборот, импульсные разряды нейронов заметно его опережают (см. рис. 6).
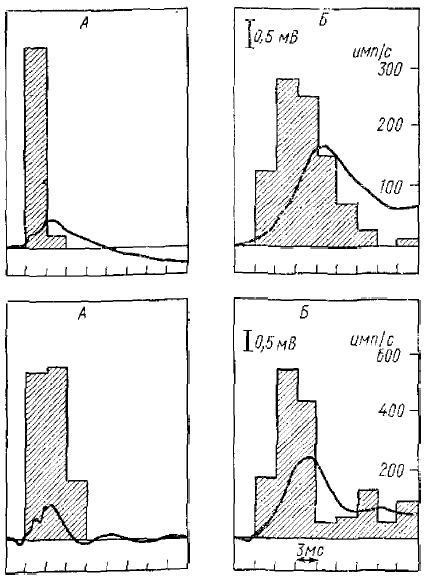
Максимум частоты импульсных разрядов наблюдается до вершины УО, а. его вершина совпадает со спадом частоты импульсных разрядов. Можно предположить, что УО является отражением постсинаптических потенциалов, вторично вызываемых импульсами, возникающими в корковых нейронах. Действительно, дальнейший анализ генеза УО, предпринятый А. Я. Супиным (1970), показал, что вспышка активности кортикальных нейронов во время УО вызвана появлением в них высокоамплитудных ВПСП, которые вызываются в свою очередь самой вспышкой импульсной активности нейронов, т. е. благодаря наличию у них возвратных возбуждающих связей. В результате такого процесса самовозбуждения и возникают высокоамплитудный ВПСП и вспышка импульсных разрядов. Таким образом, УО генерируется системой внутрикортикального возвратного возбуждения. Эти данные, полученные А. Я. Супиным, позволили объяснить и появление самого УО в момент совпадения тестирующего стимула со спадом медленной негативной волны РРП (или спонтанной α-волны).
Как связаны постсипаптическое торможение нейронов и вызванная α-подобная волна затылочной коры кролика?
Регистрация потенциалов от различных слоев коры во время α-подобной волны обнаруживает типичную картину так называемого дипольного переворота: по мере углубления регистрирующего электрода в кору негативная α-подобная волна уменьшается по амплитуде, на глубине около 0,8—0,9 мм она достигает почти нулевой амплитуды, и при дальнейшем углублении электрода вместо негативной волны начинает нарастать такой же длительности медленная позитивная волна, достигающая максимальной амплитуды в нижних слоях коры (см. рис. 4). То, что НВ сопровождается позитивной волной в глубине коры и ТПСП в корковых нейронах, дает основание для предположения, что она генерируется гиперполяризующими синапсами, расположенными на глубоких частях вертикально ориентированных (очевидно, пирамидных) нейронов коры. По-видимому, активация этих синапсов вызывает появление тока, направленного в глубине коры из клеток во внеклеточную среду, что вызывает гиперполяризацию нейронов (ТПСП) (следствием которого является торможение импульсной активности) и сдвиг экстраклеточного потенциала в глубине коры в сторону позитивности, что и регистрируется в виде медленной глубинно-позитивной волны. Поверхность коры по отношению к глубоким слоям становится негативной, что регистрируется в виде НВ. Естественно, что наиболее значимыми в решении вопроса о генезе НВ были бы внутриклеточные исследования на нейронах зрительной коры. Такие исследования пока немногочисленны, но они свидетельствуют, что в одиночных элементах зрительной области возникает длительная волна гиперполяризации как на стимуляцию зрительного нерва, так и на световые раздражения. Причем по своей длительности эта гиперполяризация хорошо коррелирует с глубинно-позитивной волной (Fuster et al., 1965; Скребицкий, Воронин, 1965; Супин, 1970). Таким образом, вполне вероятно, что гиперполяризация, лежащая в основе глубинно-позитивной медленной волны (или поверхностно-негативной волны, т. е. основного комплекса α-волны), является ТПСП.
Результаты фармакологического анализа свойств α-подобной волны, проведенные в нашей лаборатории Т. Ф. Курпяковой и А. Я. Супиным, (1967), подтвердили не только гипотезу о ее тормозном происхождении, но и позволили внести в эту гипотезу некоторые уточнения.
Было показано, что поверхностно-негативная волна является дипольным отражением активности гиперполяризующих синапсов не самых глубоких слоев коры (эти синапсы генерируют глубинно-позитивную волну), а средних слоев коры.
Ройтбак (1963, 1965) предполагает, что медленная- поверхностно-негативная волна в коре является отражением деполяризации апикальных дендритов вследствие их взаимодействия с глиальными элементами.
Выдвигается также предположение (Кондратьева и Чемоданов, 1971), что поверхностно-негативная волна отражает какой-то параллельный процесс: поляризацию апикальных дендритов и деполяризацию аксонных терминалей, ведущих к пресинаптическому торможению.
Однако существующих экспериментальных данных пока еще недостаточно, чтобы отказаться от интерпретации механизмов генеза этой волны, высказанных нами ниже.
Мы рассмотрели только основной компонент α-подобной волны — поверхностно-негативное (глубинно-позитивное) колебание. Но начальная часть волны в значительной степени формируется также поверхностно-позитивным медленным колебанием (ПВ).
Связь поверхностно-позитивного медленного колебания с активностью одиночных элементов коры
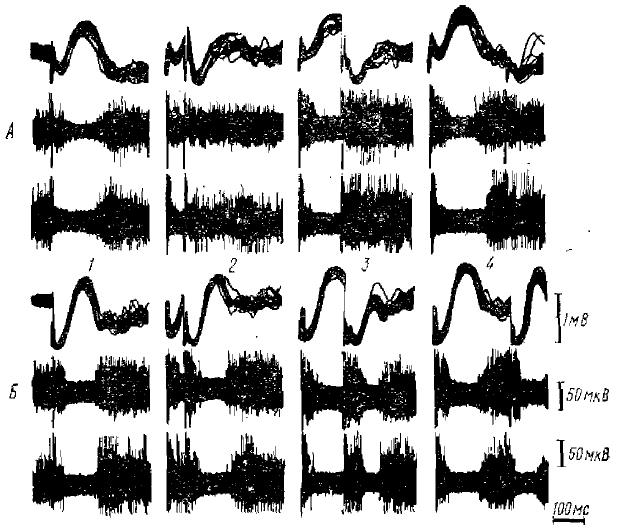
Рис. 7. Соотношение компонентов α-подобной волны — поверхностной позитивности (ПВ) и поверхностной негативности (НВ) —с импульсной активностью нейронов затылочной коры кролика (по Гусельникову и Супину, 1968)
Сопоставление ПВ с импульсной активностью нейронов различных слоев коры показывает, что в тех случаях, когда ПВ достаточно четко выражена (напомним, что она может маскироваться НВ), ее сопровождает интенсивная импульсная активность. В таких случаях импульсная реакция нейронов коры как при афферентном раздражении, так и при раздражении поверхности коры имеет следующий характер (рис. 7). Вначале наблюдается короткая (10—20 мс) вспышка активности, соответствующая по времени ПО коры (в случае афферентного раздражения) или дендритному ответу (в случае прямого раздражения коры). Затем после короткой паузы, иногда малозаметной, наблюдается повторная фаза активации, соответствующая по времени ПВ; интенсивность импульсных разрядов в это время обычно меньше, чем во время начальной активации, но все же несколько превышает фоновую активность. Длительность фазы повторной активации у некоторых нейронов довольно значительна (начало через 20—25 мс, конец, через 50 -80 не после стимула), у других нейронов очень велика, После повторной активации следует тормозная пауза, соответствующая НВ, которая заканчивается через 160—200 мс после стимула. По окончании тормозной паузы импульсная активность возобновляется, причем непосредственно вслед за паузой она может быть заметно более интенсивной, чем в фоне (явление «отдачи»), В тех случаях, когда НВ не выражена, не наблюдается повторной активации в импульсной активности нейронов, непосредственно вслед за первичной активацией наступает тормозная пауза.
Совпадение ПВ с активацией корковых нейронов позволяет считать, что происхождение ПВ связано с ВПСП в корковых нейронах. Эти ВПСП должны иметь значительную длительность (30—60 мс), по, судя по интенсивности импульсной активности, меньшую амплитуду, чем первичные ВПСП. Поверхностно-позитивная полярность волны показывает, что она может генерироваться синапсами, расположенными в относительно глубоких слоях коры. Такое предположение подтверждается анализом распределения потенциалов по поперечнику коры: при погружении регистрирующего электрода в глубину коры ПВ, так же как и НВ, претерпевает инверсию; позитивному колебанию на поверхности коры соответствует негативное колебание в глубине. Эта глубинно-негативная волна является, очевидно, прямым отражением активности глубинных деполяризующих синапсов, а ПВ является дипольным отражением активности таких синапсов.
Изложенные представления о механизмах генерации α-подобной волны вполне согласуются с имеющимися уже в литературе высказываниями о возбудительной природе ПВ (Burns, 1950, 1951; Ройтбак, 1964; Бомштейн, 1971) и тормозной природе НВ (Полянский, 1965; Кондратьева, 1964, 1967, 1969; Бомштейн, 1971).
После всего сказанного возникает вопрос, какова природа медленных ВПСП и ТПСП, возникающих во время α-подобной волны, каким путем активируются деполяризующие и гиперполяризующие синапсы корковых нейронов, активность которых приводит к возникновению позитивно-негативного (при регистрации от поверхности коры) «-подобного комплекса.
Основной поверхностно-негативный компонент α-подобной волны связан с торможением нейронной активности. Такое относительно длительное торможение, видимо, является общим свойством корковых нейронов, которое обнаруживается при реакциях коры, вызванных разными способами и в разных условиях: в зрительной коре при афферентном световом и электрическом раздражении зрительного нерва, при раздражении соматического нерва, специфических и неспецифических релейных таламических ядер и при прямом раздражении поверхности коры. Регистрация внутриклеточных потенциалов корковых нейронов показывает, что такое торможение должно являться следствием длительных и высокоамплитудных ТПСП (Phyllips, 1956, 1959; Li et al., 1960, 1962; Скребицкий, Воронин, 1966, и др.) Такая универсальность явления торможения нейронной активности наводит на мысль, что в принципе оно свойственно вообще любым видам реакций корковых, нейронов на различные раздражения, а значит, является следствием работы внутрикорковых тормозных механизмов.
По аналогии с процессами в таламических нейронах, где также наблюдается длительные торможение при различных раздражениях, можно предположить, что наблюдаемое в коре торможение является возвратным. Действительно, если существует система возвратного торможения, то она должна проявлять себя всякий раз, когда каким-либо образом вызывается первичная активация нейронов.
Принципиальная возможность существования в новой коре возвратного торможения показана вполне достоверно для нейронов пирамидного кортико-спинального тракта. Для этих же корковых нейронов показано и наличие возвратного возбуждения. Система возвратного возбуждения показана для зрительной коры кролика (Супин, 1970). Однако пока точно не известно, имеют ли такие же возвратные, но тормозные связи нейроны затылочной коры кролика, генерирующие α-подобные колебания. Аксоны этих нейронов не образуют длинных компактных трактов, которые могли бы обеспечить возможность их антидромной активации. Решить этот вопрос можно, по- видимому, другим путем.
С этой целью исследовались циклы восстановления компонентов α-подобной волны — ПВ и НВ при парных раздражениях зрительного нерва и парных раздражениях, наносимых непосредственно на кору (Гусельников, Супин, 1968). Эти данные позволили с большой степенью достоверности говорить о наличии у нейронов зрительной коры кролика системы возвратного торможения.
Однако надо отметить, что такое представление о возвратной природе торможения в зрительной коре скорее утверждается по аналогии с таковым же механизмом в других отделах мозга, где он четко показан как морфологическими, так и физиологическими исследованиями (хотя, рассуждая логически, пашей точке зрения соответствует масса фактов, приведенных выше).
Для окончательного же подтверждения этого положения, конечно, необходима точная идентификация тормозных вставочных нейронов в зрительной коре, что остается еще далеко не решенным вопросом.
Предполагается, что тормозными интернейронами зрительной коры могут являться нейроны С-группы, т. е. нейроны коры, которые, как правило, активируются во время резкого снижения активности других видов нейронов зрительной коры (В, Д, Е типа по классификации Юнга, 1964). Однако специальный анализ этого .положения (Хаютин, 1973) не дает оснований считать С-нейроны специфическими тормозными вставочными нейронами.
Таким образом, изложенные результаты исследований позволяют представить следующую последовательность процессов в нейронах затылочной коры кролика после одиночного афферентного раздражения или прямого раздражения коры. Активация возбуждающих синапсов корковых нейронов, отражающаяся в появлении глубинно-негативной фазы ПО (при афферентном раздражении) или дендритного ответа (при прямом раздражении коры) вызывает разряды пирамидных нейронов. Через систему возвратных связей (коллатерали аксонов или восходящие аксоны и, вероятно, вставочные нейроны) эти импульсы (а в случае раздражения коры также и импульсы, возникшие в результате непосредственного раздражения аксонов и вставочных нейронов) возвращаются к пирамидным нейронам, вызывая в них сначала ВПСП (с ними связана ПВ) и еще более длительные ТПСП (с ними связана НВ). Тормозные синапсы создают более мощные токи, чем возбуждающие, в результате этого активность возбуждающих синапсов может маскироваться активностью тормозных синапсов, что проявляется как отсутствие поверхностно-позитивной волны, которое может наблюдаться при афферентных раздражениях.
Представление о возвратной природе активации синапсов, генерирующих α-подобную волну, легко объясняет повышение возбудимости нейронов коры, наблюдаемое в конце поверхностно-негативной волны. Фоновые разряды нейронов благодаря наличию системы возвратного торможения приводят к постоянной фоновой активации тормозных синапсов, что ограничивает частоту импульсных разрядов нейрона на определенном уровне — возникает своего рода стабилизирующая система с отрицательной обратной связью. Раздражение, вызывающее интенсивный разряд большого количества нейронов, приводит к тому, что фоновая активность тормозных синапсов сменяется ТПСП, возникшим в результате синхронной возвратной активации сразу большого количества тормозных синапсов. Большая величина этого ТПСП приводит уже не к ограничению частоты, а к полному подавлению импульсных разрядов. В связи с отсутствием импульсных разрядов нейронов активность тормозных синапсов больше не поддерживается — ТПСП спадает с определенной скоростью, зависящей, возможно, от скорости рассасывания или разложения медиатора или от других причин. Что же происходит, когда внутриклеточный потенциал нейрона уменьшится до величины, при которой возобновятся импульсные разряды? Система возвратного торможения не может обеспечить мгновенное ограничение частоты импульсных разрядов, так как возвратная активация тормозных синапсов имеет некоторый латентный период, да и по окончании этого латентного периода тормозной эффект, очевидно, не мгновенно достигает максимальной величины, а развивается в течение некоторого промежутка времени. Все это время продолжается уменьшение внутриклеточного потенциала нейронов и нарастание частоты их импульсной активности, которая может достигнуть значительно большей величины, чем в фоне. Нарастание деполяризации и усиление импульсной активности нейронов может значительно ускоряться действием системы возвратного возбуждения, которая, судя по соотношению временных параметров поверхностно-позитивной и поверхностно-негативной волн, является более быстродействующей, чем система возвратного торможения. Деполяризация нейронов приводит также к тому, что ВПСП, вызываемые на этом фоне афферентными раздражениями, дают наибольший эффект — максимальные импульсные реакции и максимальную амплитуду УО. Только по истечении времени, необходимого для развития эффекта возвратного торможения, внутриклеточный потенциал и частота импульсной активности нейронов могут вновь стабилизироваться на исходном уровне.
Предлагаемая интерпретация последовательности явлений хорошо объясняет и происхождение позднего вызванного потенциала затылочной коры кролика (экстрапотенциала). Ранее было показано, что разряды корковых нейронов, возникшие в ответ на афферентное раздражение, сопровождаются вызванным потенциалом — УО. Аналогичным потенциалом должна сопровождаться и вспышка импульсной активности, появляющаяся по окончании α-подобной волны, — возникает поздний вызванный потенциал. Вполне естественно, что вспышка активности нейронов и поздний вызванный потенциал должны наблюдаться после каждой α-подобной волны.
Механизмы генерации спонтанной α-подобной активности
Регистрация спонтанной активности одновременно от поверхности и из глубины коры обнаруживает характерную картину синхронных колебаний противоположной полярности на поверхности и в глубине при появлении ритмической активности α-подобного типа. Микроэлектродная регистрация импульсной активности нейронов коры показывает, что каждой НВ (глубинно-позитивной) волне соответствует более или менее глубокое торможение спонтанной импульсной активности, а каждой ПВ (глубинно-негативной) — вспышка импульсной активности. Очевидно, этот факт полностью соответствует представлению, согласно которому спонтанная α-подобная активность, как и вызванные α-подобные волны, отражает деятельность преимущественно тормозных синапсов, генерирующих основной компонент активности — НВ (глубинно-позитивные) волны, а также в некоторой степени возбуждающих синапсов, генерирующих ПВ (глубинно-негативные).
Итак, генераторами всех видов α-подобной активности можно считать возбуждающие и тормозящие синапсы (преимущественно тормозящие), расположенные в различных слоях коры и активируемые через посредство возвратных связей, причем активность, регистрируемая от поверхности коры, является отражением деятельности не поверхностных, а относительно глубоко расположенных синапсов.
Синапсы глубоких слоев коры, в первую очередь тормозящие, могут играть важную роль в генерации не только α-подобной активности, но и других видов спонтанной электрической активности коры. Так, длительные ТПСП появляются в нейронах коры во время реакции вовлечения у слабо наркотизированных животных. Кроме того, рядом авторов отмечено, что нерегулярная, аритмичная спонтанная электрическая активность коры сопровождается учащением импульсных разрядов нейронов при сдвиге потенциала поверхности коры в сторону позитивности и угнетением импульсных разрядов при сдвиге поверхностного потенциала в сторону негативности (I. Calvet, С. Calvet, 1963: Fromm, Bond, 1964; Fuster et al., 1965; Robertson, 1965, и др.). Активная тормозная природа отрицательной фазы δ-волны при одновременной регистрации нейронов коры у кролика выявилась в опытах Гринявичуса (1971). Джаспером и Стефанисом (Jasper, Stefanis, 1965) при внутриклеточных исследованиях было показано, что медленные поверхностно-негативные потенциалы могут соответствовать гиперполяризации нейронов, а поверхностно-позитивные — деполяризации нейронов. Это указывает на то, что в этих случаях электрическая активность генерируется глубоко расположенными синапсами, так как в противном случае при поверхностном расположении синапсов, генерирующих суммарную активность, соотношение потенциалов поверхности коры с активностью нейронов должно было бы быть противоположным (негативность при деполяризации и позитивность при гиперполяризации). Наличие глубокого угнетения импульсной активности во время поверхностно-негативных волн показывает, что весьма возможно участие в этом процессе тормозных, гиперполяризирующих синапсов. Не исключено также, что такие формы нерегулярной электрической активности коры родственны α-подобной активности. Основное отличие между ними может состоять в том, что в одном случае активность синапсов в значительной степени асинхронна и более или менее четкое соотношение суммарной активности и активности одиночных нейронов наблюдается относительно редко, когда однотипные процессы в большинстве нейронов совпадут. В другом же случае имеет место длительная синхронность работы отдельных элементов, в которых ритмически чередуются фазы возбуждения и торможения. Синхронность работы большого количества элементов и приводит к появлению ритмических суммарных колебаний потенциала в коре, имеющих значительную амплитуду и обнаруживающих корреляцию с активностью большинства нейронов.
До сих пор мы разбирали только α-подобную активность животных. Но, безусловно, значительный интерес представляет также вопрос о том, имеет ли такие же механизмы и α-ритм человека.

Интересны в этом отношении данные о циклах возбудимости корковых элементов, сопровождающих циклы α-активности. Еще Лэнсингом (Lansing, 1957) было установлено, что латентный период, двигательной реакции человека (движение, которое испытуемый должен совершать при предъявлении световой вспышки) зависит от фазы волны α-ритма (рис. 8), с которой совпадает световой стимул: наиболее короткий латентный период (рис. 8, А, непрерывная линия) имеют реакции на вспышки света, даваемые в моменты около негативной вершины α-волны, наиболее длинный (пунктирная линия) латентный период наблюдается, если вспышки даются около позитивной вершины волны. Более того, сами двигательные реакции не одинаково вероятно совпадают с различными фазами α-волн: чаще всего реакции наблюдаются в моменты около позитивной вершины (рис. 8, В, непрерывная линия), реже — около негативной (пунктирная линия) вершины. Если ввести поправку на время проведения возбуждения от сетчатки до зрительной коры (около 30 мс) и на время проведения возбуждения от двигательной коры к мышцам (около 15 мс), то получается (рис. 8, Б, Г), что по обоим признакам (минимальный латентный период реакции и максимальная вероятность появления реакции) максимальная возбудимость элементов коры должна совпадать с одной и той же фазой α-волн — с переходом негативной вершины к позитивной. Минимальная возбудимость — с переходом от позитивной вершины к негативной (на Б и Г — пунктирная линия).
Это наблюдение в дальнейшем детально разработали Кэллуей и соавторы (Callaway et al., 1960, 1961, 1962). Более того, максимальная возбудимость коры в моменты, соответствующие переходу от негативных вершин α-волн к позитивным, определяется также по максимальной амплитуде вызванных потенциалов на световые раздражения (Callaway, Layne, 1964; Dustman, Beck, 1965). Корреляцию уровня возбудимости с фазами α-волн у человека нашли и ряд других исследователей (Асафов, 1971 и др.).
Эти результаты полностью соответствуют данным об изменении возбудимости корковых нейронов во время α-подобной волны у кролика, по крайней мере, они не противоречат предположению об α-волнах человека, как отражений чередования генерируемых в глубине коры ТПСП (поверхностно-негативные волны) и ВПСП (поверхностно-позитивные вершины).
Циклы восстановления α-волн человека также аналогичны циклам восстановления α-подобных волн коры мозга кролика. На это указывают частотно-амплитудная характеристика реакций перестройки ритма. Реакция перестройки ритма у человека (см. лекцию 13) имеет максимальную амплитуду при частоте стимуляции около 10 имп/с, когда каждый последующий залп импульсов из сетчатки совпадает с концом α-волны, вызванной предыдущим стимулом. Учащение стимуляция ведет к сильному уменьшению амплитуды вызванных α-волн (реакции перестройки), что вполне согласуется с представлением о тормозной природе поверхностно-негативной α-волны. Аналогичные данные могут быть обнаружены и при использовании триггерной световой стимуляции (Бехтерева, Зонтов, 1961, 1962).
Таким образом, колебания возбудимости, совпадающие с каждым α-циклом, влияют и на амплитуду самих же вызванных α-волн. Этот вывод находится в полном соответствии с предположением, что процессы, отражающиеся в α-ритме коры мозга человека, имеют, как и при α-подобном ритме коры мозга кролика, возвратную природу (возвратное возбуждение и возвратное торможение).
Рассмотренные характеристики α-ритма человека не противоречат допущению, что он имеет такие же механизмы, как и исследованная нами α-подобная активность затылочной коры мозга кролика. Это не доказывает, что эти виды активности имеют одинаковые механизмы, но аналогичность их представляется вполне возможной.
Вернемся теперь к вопросу о РРП. Резюмируя все сказанное, можно полагать, что существует прямая аналогия между РРП и определенными видами фоновой биоэлектрической активности. Не исключено, что РРП, регистрируемый у животных в таламусе и коре, является модификациями либо α-подобной активности, либо α-активности.
Что касается РРП и α-подобной активности, то этот вопрос на примере зрительной коры кролика мы рассмотрели подробно.
При определенной стадии естественного или барбитурового сна как в таламусе, так и различных зонах коры и некоторых других структурах мозга при действии различных раздражений возникают характерные РРП, внешне напоминающие вспышку веретена σ-активности. Как мы покажем дальше (см. лекцию 9), эти РПП есть не что иное, как проявление активации механизмов, генерирующих спонтанный ритм.
РРП в других структурах, кроме коры и таламуса, изучены слабо. Однако показано, что генерация РРП, аналогичных стресс-ритму и обонятельному ритму, в принципе возможна. Одиночное электрическое раздражение первичного очага стресс-активности — перегородки — может вызвать в гиппокампе вспышку затухающей ритмической активности, по частоте и форме вполне сходной со стресс-ритмом, возникающим при тонической активации перегородки через посредство активирующих стволовых структур (Briicke et al., 1959). Очевидно, такую вспышку активности вполне можно рассматривать как родственный θ-ритму РРП (см. лекцию 10).
В нормальных условиях РРП, аналогичные ритмической активности обонятельного анализатора, были получены на лягушке в пашей лаборатории. При сильном электрическом раздражении обонятельного нерва в обонятельной луковице вслед за вызванным потенциалом на его позитивной фазе регистрируется серия ритмических колебаний или РРП (см. лекцию 11).
Анализ происхождения этого РРП в обонятельной луковице показал полную его идентичность с ритмической активностью обонятельной луковицы.
Таким образом, все сказанное еще раз подчеркивает, что способность к генерации РРП не является качественным, принципиальным отличием генераторов ритмической активности в коре и таламусе от других генераторов ритмической активности.
Роль подкорковых структур в происхождении α-ритма человека и α-подобной активности животных
Все рассматриваемые виды ритмической активности не возникают в какой-либо одной структуре головного мозга, а регистрируются одновременно, по крайней мере, от нескольких структур, объединенных взаимными связями. Ритмическая активность возникает в них одновременно, имеет обычно одинаковую частоту и постоянные фазовые соотношения. Это дает основание предполагать, что, по крайней мере, в некоторых отделах головного мозга ритмическая активность может генерироваться не самостоятельно, а под влиянием ритмических залпов импульсов из других образований, где эта активность первично возникает. Структуры, являющиеся первичными очагами ритмической активности, часто называют водителями (pacemaker), поскольку они задают ритм другим структурам.
Возникает вопрос, не является ли и α-ритм коры мозга человека результатом работы подкоркового водителя. Этот вопрос до сих пор остается нерешенным.
В связи с этим определенный интерес представляют данные клиники об изменениях электрической активности коры мозга при различных поверхностных и глубинных поражениях (Бехтерева, I960). Поражения многих подкорковых структур мозга сопровождаются весьма заметными изменениями электрической активности коры, в том числе изменениями α-ритма. Однако пока мало фактов, которые .могли бы помочь решить вопрос о том, являются ли эти изменения результатом воздействия на подкорковый водитель α-ритма или же они наступают вследствие изменения функционального состояния коры при выключении подкорковой афферентации.
Другой метод анализа, который применялся с целью решения вопроса о подкорковом первичном очаге α-активности — исследование синхронности α-ритма в различных областях коры. Полученные результаты Гароутта и соавторов (Garoutte et al., 1956, 1958, 1959), показали, что внутриполушарная и особенно межполушарная синхронность α-колебаний довольно высока, что было расценено авторами как доказательство существования подкоркового водителя α-ритма, контролирующего синхронную активность всех точек коры. Однако авторы не приводят убедительных доказательств того, что в синхронизации α-активности между различными точками коры не могут играть основной роли кортико-кортикальные связи. Так что и этот метод пока не дал определенных результатов и вопрос о первичном очаге α-ритма остался открытым.
Оригинальную гипотезу о подкорковом водителе α-ритма у человека выдвинули Кандел и Энгелей (Keidel, Engelen, 1971). Они полагают, что в области коленчатых тел имеется «водитель ритма», который одновременно задает частоту α-ритма в затылочных отделах мозга и частоту иннервации глазных мышц. Однако эта гипотеза требует также экспериментального подтверждения.
Что касается α-подобного ритма зрительной коры кролика, то возникает вопрос — может ли система возвратного торможения обеспечить автономное возникновение в коре ритмической активности за счет тормозной синхронизации корковых нейронов.
Все данные, свидетельствующие о первично-таламическом происхождении ритмической активности коры, относятся только к одному определенному виду ритмической активности — веретенообразной активности мозга (σ-ритму). Поскольку веретенообразная активность мозга животных является хорошо изученным видом ритмической активности, часто возникает тенденция относить данные, полученные при ее изучении, вообще к любой ритмической активности коры и таламуса, в том числе к α-ритму (Jasper, 1949; Bremer, 1955, 1958; Andersen, Andersson, 1968). Однако такая тенденция едва ли оправданна (Elul, 1972). Как раз на примере ЭЭГ человека видно, что α-ритм и сонные веретена представляют собой различные ЭЭГ-феномены. не зависящие друг от друга и имеющие различные свойства. α-подобные колебания у животных также имеют несколько иные свойства, чем веретенообразная активность. Это заставляет с большой осторожностью отнестись к мнению, что все виды ритмической активности коры являются следствием только таламической ритмической активности.
Далее, опыты показывают, что в некоторых условиях изолированный участок коры может генерировать ритмическую активность. В этом случае ритмическая активность наблюдается при химической стимуляции ацетилхолином в комбинации с антихолинэстеразными препаратами (эзерином и др.) (Jasper, 1949; Gol, Callaway, 1963). Кроме того, примером ритмической активности изолированной коры может служить так называемый судорожный разряд последействия — длительная вспышка весьма регулярной, ритмической активности, возникающая в изолированном участке коры после тетанизации и длящаяся до нескольких минут (Burns, 1950, 1951; Echiin et al., 1952, и др.), причем легче, чем в интактной коре, хотя эти виды ритмической активности можно рассматривать как патологические.
Такая ритмическая активность изолированной коры может представить определенный интерес, если раскрыть механизмы ее генерации и проверить применимость этих механизмов к нормальной спонтанной ритмической активности интактной коры.
Говоря о возможности применения гипотезы тормозной синхронизации к нейронам коры мозга, прежде всего обращаешь внимание на α-подобную активность коры. Мы показали, что α-подобным волнам соответствует появление длительных ТПСП в нейронах, имеющих возвратную природу. Можно предположить, что ритмическая последовательность таких ТПСП может возникать в коре автономно, без синхронизирующих влияний из таламуса, т. е. разряд группы нейронов коры может вызвать в них длительный (около 200 мс) ТПСП, который затормозит дальнейшие разряды, а после окончания ТПСП разряды нейронов возобновятся и усилятся благодаря действию системы возвратного возбуждения, в результате чего через систему возвратных тормозных связей будет вызван новый ТПСП, опять прерывающий разряды нейронов на период около 200 мс и т. д. В результате возникает ритмическая активность нейронов с периодом около 200 мс, т. е. с частотой около 5 Гц; именно такую частоту и имеет α-подобная активность затылочной коры кролика. Синхронность активности отдельных нейронов обеспечивается перекрестным характером тормозных связей. Причем ритмическая активность затылочной коры кролика отнюдь не является исключением.
Данные, указывающие на тормозную ритмическую синхронизацию активности корковых нейронов, были получены также на соматосенсорной коре кошки Андерсоном (Andersson, 1965). Он показал, что при внутриклеточной регистрации активности корковых нейронов можно выделить два типа ритмических разрядов последействия: первый имеет первично-таламическое происхождение, а второй обусловлен внутрикортикальной тормозной синхронизацией нейронной активности. Однако пока точно не известно, действительно ли для образования каждой последующей волны α-подобной активности (т. е. для вызова возвратных ТПСП в нейронах коры) достаточно разрядов корковых нейронов, возникающих после предшествующего ТПСП, или же для запуска каждой новой волны α-подобной активности необходим добавочный синхронизирующий залп импульсов из таламуса.
В связи с этим как раз и представляет интерес сравнение α-подобной активности с ритмической электрической активностью коры, изолированной от таламуса.
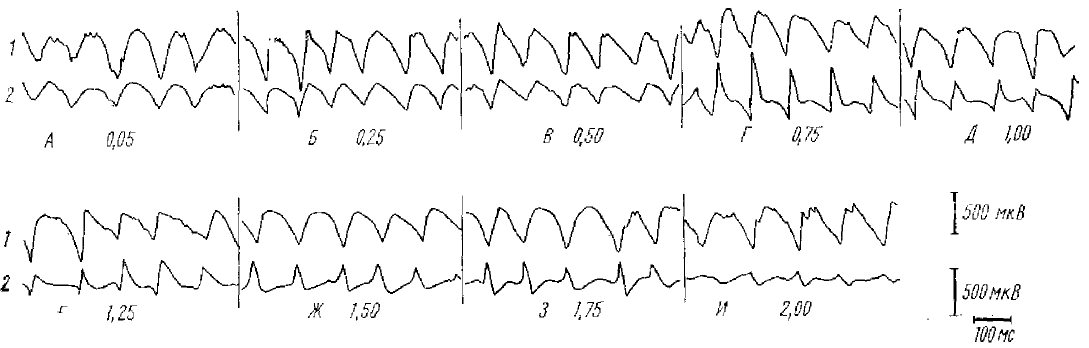
Исследование судорожного разряда последействия в коре изолированного полушария (Супин, 1967) показывает (рис. 9), что он во многих отношениях очень сходен с α-подобной активностью нормальной коры.
Во-первых, он имеет строго ритмичный регулярный характер и состоит из медленных поверхностно-негативных волн (сходных с НВ α-подобного ритма), чередующихся с более быстрыми пикоподобными компонентами преимущественно позитивной полярности (сходными с поздними вызванными потенциалами нормальной коры, которые возникают после каждой α-подобной волны).
Во-вторых, при погружении регистрирующего электрода в глубь коры медленные волны и пикоподобные компоненты судорожного разряда последействия претерпевают реверсию. Медленные волны в глубине коры позитивны, а пикоподобные компоненты — негативны. Реверсия медленных волн проходит через стадию почти нулевой амплитуды (глубина 1,00—1,25 мм). Реверсия медленных волн происходит на несколько большой глубине, чем реверсия пикоподобных компонентов: при погружении электрода на глубину, где пикоподобные колебания уже полностью инвертированы, медленные волны еще могут иметь ту же полярность, что и на поверхности (глубина 0,75 мм). Все эти характеристики послойного распределения потенциалов медленных волн и пикоподобных компонентов полностью совпадают с картиной послойного распределения α-подобной активности (медленных α-подобных волн и поздних вызванных потенциалов) .
В-третьих, суммарные потенциалы коры во время судорожного разряда последействия имеют точно такое же соотношение с импульсной активностью корковых нейронов, как и суммарные потенциалы α-подобной активности. Во время каждого поверхностно-позитивного (глубинно-негативного) пика судорожного разряда наблюдается четко выраженная относительно короткая (20— 50 мсек) вспышка активности. Каждой поверхностно-негативной (глубинно-позитивной) волне соответствует период полного молчания нейронов.
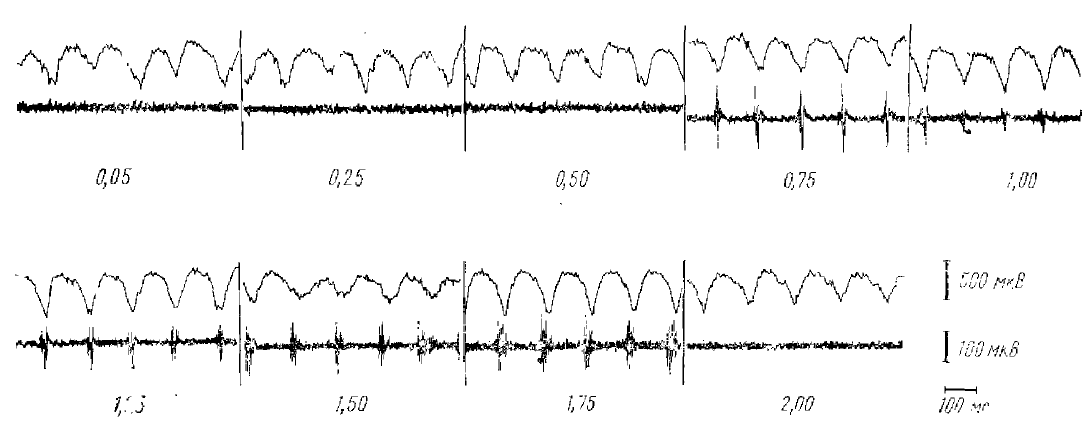
Переход от вспышки активности к тормозной паузе происходит не постепенно, а в виде резкого прекращения активности. Такая картина наблюдается в коре на любой глубине где только могут быть зарегистрированы импульсные разряды (рис. 10). Если сопоставить эту картину с картиной, описанной для α-подобной активности, то видно их полное соответствие.
Вее эти характеристики судорожного разряда последействия, а также его сходство с α-подобной активностью изолированной коры дают некоторое основание предположить (Супин, 1967), что медленные поверхностно-негативные волны разряда являются отражением длительных ТПСП, блокирующих активность корковых нейронов. Эти ТПСП являются, очевидно, возвратными: на это указывает как сходство их с возвратными ТПСП α-подобной активности, так и тот факт что они возникают в деафферентированной коре, т. е. должны вызываться разрядами самих же корковых нейронов. Пикоподобные компоненты, связанные со вспышками импульсных разрядов, являются, очевидно, отражением ВПСП. Такие ВПСП в нейронах деафферентированной коры могут возникать в результате активности возвратных возбуждающих связей.
Итак, системы возвратного торможения и возвратного возбуждения принимают, по-видимому, самое непосредственное участие в генерации судорожного разряда последействия. Но, поскольку этот вид ритмической активности может возникать в изолированной коре (отделенной от таламуса), можно допустить, что тормозная синхронизация активности нейронов на корковом уровне в принципе возможна, по крайней мере, для одного вида ритмической активности коры — судорожного разряда последействия.
Некоторое сходство между судорожным разрядом последействия и α-подобной активностью коры дает возможность предполагать, что и α-подобная активность может иметь тот же самый механизм. Такую же общность генеза α-ритма и судорожного РРП отмечают Р. И. Кругликов и др. (1970). Эти результаты имеют прямое отношение к вопросу о первичном очаге α-подобной активности. Они показывают, что, возможно, первичным очагом может быть сама кора.
Вопрос, о механизмах α-ритма человека остается неясным. Возможно, что он, как и α-подобный ритм коры мозга кролика, генерируется в результате тормозной синхронизации активности корковых нейронов. Но такое предположение носит, конечно, чисто гипотетический характер. Не исключено, что α-ритм человека— это совершенно новый вид активности, который не имеет ничего общего с α-подобным ритмом кролика.
В связи с этим вызывает интерес сообщение Куло и др. (Kuhlo et al., 1968) об α-подобном ритме ЭЭГ человека, частотой 4—5 в 1 с, возникающего при некоторых патологических отклонениях. Этот ритм близок по своим параметрам к α-подобному ритму кролика и α-ритму человека, хотя его происхождение далеко не ясно.
К настоящему времени наши знания о генезе α-ритма человека совершенно недостаточны для каких-либо заключений. Более того, отсутствие этих знаний даже привело ряд исследователей к мнению об экстрацеребральном, не мозговом происхождении α-ритма человека, что безусловно не оправдано (Butler, Glass, 1970; Chapman ct al., 1971; Lippold et al., 1970, 1971).
Так, например, Липольд (1970) считает, что -ритм генерируется не в мозговой ткани, а является результатом распространения на череп постоянного потенциала глазного яблока, колебательный характер этого потенциала связан с его модуляцией тремором глазодвигательных мышц.
И если нет оснований, с нашей точки зрения, сомневаться в истинности α-активности ЭЭГ человека, как физиологического явления, то более странным является то, как отметил Блуд (Elul, 1972), что эта проблема, которая с большим приближением могла бы быть решена на приматах (так как последние обладают выраженным α-ритмом), пока интенсивно не разрабатывается.