Электрическую активность головного мозга в виде электроэнцефалограммы (ЭЭГ) человека впервые зарегистрировал австрийский психиатр Ганс Бергер в 1924 г. В начале 30-х гг. были опубликованы результаты его многолетних исследований. К настоящему времени в намеченных им областях применения электроэнцефалографии – клинике, фармакологии, физиологии и психологии, – благодаря усилиям нескольких поколений исследователей накоплен огромный экспериментально-клинический материал, изложенный и частично обобщенный в многочисленных статьях, монографиях, методических руководствах.
Поскольку феноменологическое описание ЭЭГ повторяется во многих работах, нет необходимости цитировать множество литературных источников, вполне допустимо ограничиться наиболее важными, с нашей точки зрения, т. е. для системного анализа ЭЭГ.
Прежде, чем говорить о системном анализе ритмов электрической активности головного мозга целесообразно хотя бы коротко сказать о предмете системного анализа. Несмотря на огромное количество литературы по энцефалографическим исследованиям многие вопросы механизма возникновения ритмики осцилляторных процессов головного мозга до сих пор остаются открытыми.
О механизмах формирования ритмики электроэнцефалограммы
Прежде всего следует отметить, что содержание приведенного ниже краткого обзора ограничивается следующими условиями. Во-первых, в понятие «ритм» электроэнцефалограммы мы вкладываем содержание, соответствующее принятому в физике термину «осцилляторный случайный процесс». Во-вторых, отметим, что до настоящего времени еще отсутствуют общепринятые представления о природе формирования ЭЭГ и физических механизмах, регистрируемых в мозге и окружающих его тканях суммарных биопотенциалах1. Уделим внимание основным модельным представлениям формирования ЭЭГ и методам ее анализа.
Первая модель генерации ЭЭГ, появившаяся в 30-х гг., рассматривала электроэнцефалограмму как сумму импульсных разрядов клеток коры головного мозга [R. Adrian, Matthews, 1935]. О широком признании указанной точки зрения свидетельствуют работы2. Именно в то время были введены понятия «синхронизация» и «десинхронизация»[R. Adrian, Matthews, 1935]. Появление в электроэнцефалограмме высокоамплитудных медленных волн приписывали процессу слаженной деятельности корковых элементов, в то время как низкоамплитудные быстрые колебания связывали с некоторым нарушением этой согласованности. Развитие микроэлектродной техники в 50-х гг. способствовало получению экспериментальных фактов, противоречащих модели Эдриана и Мэтьюса. Было показано, что медленная активность не распадается на спайки – они аддитивно накладываются на нее3. При этом спонтанная импульсная активность корковых нейронов почти не связана с ЭЭГ 4.
Отсутствие указанной связи проявляется не только во времени, но и в пространстве: импульсная активность локализована в коре с высокой степенью точности). Суммарная активность не имеет аналогичного фокуса5. К сказанному можно добавить, что в состоянии сна, когда количество разряжающихся нейронов существенно уменьшается, в ЭЭГ появляются высокоамплитудные медленные волны67.
Следующий шаг на пути исследования природы ЭЭГ связан с использованием большого числа близко расположенных микроэлектродов. Непрерывная запись электрической активности с помощью микроэлектродов наглядно продемонстрировала, что источником ЭЭГ является мембрана нейрона8. Одновременное визуальное наблюдение внутриклеточной активности и суммарной ЭЭГ, а также кросс-корреляционный анализ 10-секундных отрезков записи9 привели к представлению о взаимной независимости элементарных источников электроэнцефалограммы. Позднее М. Н. Жадиным было показано10 на основании использования аппарата теории поля и математической статистики, что элементарным генератором следует считать клеточный диполь единичного корневого нейрона, возникающий в результате пространственно-временной суммации постсинаптических потенциалов этой клетки, вызываемых импульсной активностью множества других клеток.
Дипольные моменты таких полей обусловлены асимметрией ветвления клетки. Их количественная оценка дана в работах11 [А. М. Гутман, 1980]. В рамках указанной модели ЭЭГ является результатом слабо скоррелированной активности одинаково ориентированных пирамидных клеток коры [М. Н. Жадин, 1969]. Показано12, что равномерное распределение коэффициента корреляции между парами диполей приводит к мощному эффекту синхронизации. Зависимость же коэффициента корреляции ЭЭГ от расстояния между электродами может быть обусловлена повышенным уровнем корреляции между близко расположенными нейронами. Как отмечается в работе [М. Н. Жадин, 1981], клеточный диполь – не единственный возможный элементарный источник ЭЭГ. Весьма естественным представляется введение в качестве такого источника понятия «кванта ЭЭГ» [Ю. Груодис, А. М. Гутман и др.,1972]. По определению авторов, «квант ЭЭГ» – это внеклеточное отражение большого числа индивидуальных постсинаптических потенциалов, синхронно возникающих в разных клетках в ответ на один афферентный спайк. Однако такое определение кванта ЭЭГ весьма неопределенно и не соответствует кванту физике осцилляторного процесса. В работе [М. Н. Жадин, 1981] показано преимущество подхода на основе представления о клеточном диполе, которое состоит в том, что средние величины коэффициентов корреляции между импульсными потоками, генерируемыми клетками – параметры, доступные непосредственному экспериментальному измерению.
Таким образом, модель М. Н. Жадина дает представление о связи ЭЭГ с элементарным источником ЭЭГ (клеточным диполем). За рамками модели остаются вопросы формирования ритмики в составе ЭЭГ и ЭкоГ, так как при этом на первый план выступают системные свойства мозга. Это означает, что составляющие системы, не обязаны обладать свойством, присущим системе в целом. [Babloyantz et al., 1986].
Такой подход к ЭЭГ можно рассматривать как системный. Надо сказать, что основное внимание большинства исследователей на протяжении всей истории развития метода электроэнцефалографии уделялось изучению ритмических процессов, поскольку они представлялись наглядным проявлением самоорганизации мозговых явлений. Представления такого рода и лежат в основе большинства современных разработок, относящихся к механизмам формирования электроэнцефалограммы. Все их многообразие может быть систематизировано в рамках элементарной классификации, определяемой идеологией общей теории колебаний. Поэтому прежде всего был проведен поиск процессов, характерные периоды которых были бы соизмеримы с периодами колебаний основных ритмов ЭЭГ (порядок – 0,1 с)13 [R. Lorente de No, 1935]. Было выявлено существование достаточно длинных цепей из последовательно включенных нейронов, синаптическая задержка каждого из которых составляет величину порядка 1 мс и автоколебаний в виде устойчивых предельных циклов.
Такой подход оказался весьма плодотворным, поскольку позволил широко использовать ЭВМ для «проигрывания» различных ситуаций, связанных с нейронными сетями (см. выше). Вопрос о реальном положении дел остается открытым.
Другим представлением о ритмической активности ЭЭГ является представление об автогенераторах, существующих либо в самой структуре, откуда регистрируется ЭЭГ, либо в другой, способной к авторитмичности («пейсмекер») и связанной с первой проводящими путями мозга. Представление об ЭЭГ-ритмах человека как результате синхронизации ряда автогенераторов, имеющих близкие частоты, принадлежит Норберту Винеру14. Авторы работы [F. Н. Lopes de Silva et al., 1973] пришли к выводу вывод о том, что спонтанный ритм у не-наркотизированного животного не может быть объяснен влиянием на кору большого количества таламических пейсмекеров. Однако более убедительными являются результаты, полученные [Р. Andersson et al., 1971] которые показали, что происхождение альфа-ритма может быть обусловлено импульсной активностью со стороны зрительного бугра. Это хорошо согласуется с клиникой – при поражении зрительного бугра наблюдается различного рода изменения альфа-активности. Е. А. Жирмунская полагает, что частота альфа-активности и ее регулярность определяется сбалансированным влиянием на кору систем регуляции, составляющих неспецифический лимбико-ретикулярный комплекс.
Как усиление, так и ослабление регулирующих посылок вызывает нерегулярность (разброс частоты) альфа-ритма. Еще более сложным представляется генез бета-активности, которая для здорового человека вообще не характерна15. Напротив, тэта-активность с небольшим индексом многие клиницисты полагают характерной для здорового человека. При значительном увеличении индекса многие клиницисты связывают его с психической патологией личности. Аналогично рассматривается и дельта-активность. Ограничимся этими замечаниями относительно клинического значения основных ритмов ЭЭГ, так как нашей задачей не является рассмотрение клинического аспекта ЭЭГ.
Нетрудно заметить, что общий недостаток описанных моделей -отсутствие экспериментальных доказательств их адекватности реально существующей динамической системе. Все эти модели имеют слишком общий характер и, главное, оставляют неопределенным вопрос о физическом смысле измеряемых характеристик колебательных составляющих ЭЭГ и функциональном порядке ритмов, и соответственно, об их физиологической (клинической) интерпретации. Эти вопросы представляются важнейшими, с нашей точки зрения.
Можно отметить повышенный интерес к вопросам мониторинга и использования автоматизированных экспертных систем, например, В. К. Davey; W. R. Fright; F. J. Caroll; R. D. Jones16. Особого внимания заслуживает рост числа исследований, связанных с так вызываемым картированием мозга (последнее обусловлено скорее всего определенными удобствами многоканальной визуализации ЭЭГ-наблюдений за счет использования дисплейной техники). Отсутствие отличающихся существенной новизной результатов, полученных подобным образом, обусловлено тем обстоятельством, что в принципиальном плане использование этой техники не содержит ничего нового по сравнению с обычным визуальным наблюдением.
Продолжает развиваться направление, связанное с внедрением в теоретическую нейробиологию представлений синергологии (самоорганизация, аттракторы и т. д.). Определенная сдержанность, которая проявляется при оценке полученных результатов и перспектив такого подхода, связана, по-видимому, с трудностями интерпретации таких результатов.
Современные методы исследования ЭЭГ
Являясь до последнего времени единственным неинвазивным методом исследования деятельности головного мозга, метод энцефалографии постоянно совершенствуется. Возрастает уровень его технической оснащенности и экспериментально-теоретической обоснованности. Расширяется число математических методов первичной компьютерной обработки ЭЭГ. Разработанный на основе ЭЭГ метод вызванных потенциалов нашел свою собственную область широкого применения в клинической диагностике и экспериментальных исследованиях. Словом, потенциальные возможности метода энцефалографии далеко еще не исчерпаны.
В разработке методов анализа ЭЭГ можно обозначить два основных направления: клиническое и экспериментально-математическое.
В клинических исследованиях выделены и систематизированы особенности ЭЭГ при эпилепсии, нарушениях мозгового кровообращения, травмах головного мозга и некоторых других заболеваниях17. Электроэнцефалографические обследования в клинике стали повседневной практикой. В последний годы многих исследователей привлекает «континиум» функциональных состояний, поскольку здесь скрыты механизмы дезадаптации, приводящей к патологии. Их раскрытие с использованием метода энцефалографии встречает две фундаментальные проблемы.
Первая – недостаточность всех способов первичной обработки ЭЭГ, обусловленная тем, что ЭЭГ представляет собой нестационарный, нелинейный, случайный многомерный процесс. Анализ таких процессов относится к числу еще нерешенных проблем, и для ее решения широко используется подход, который рассматривает ЭЭГ на коротких интервалах в качестве эргодического процесса. Такой подход позволяет использовать автоматизирование анализа ЭЭГ с использованием хорошо разработанных методов физико-математического анализа эргодических процессов.
Другая трудность – отсутствие общепринятых представлений о физиологической и патофизиологической значимости элементов и отдельных паттернов ЭЭГ. Ее первопричина определяется сложностью структурно-функциональных механизмов генерации ЭЭГ. Очевидно, в результате преодоления этих трудностей содержательный анализ ЭЭГ достигнет такой же высокой степени, которая имеется, например, в электрокардиографии. Однако уже теперь метод ЭЭГ позволяет ставить новые научно-практические вопросы, сама постановка которых без него была бы невозможна.
Обзор литературы по этой теме сделан исчерпывающе А. П. Кулаичевым18. В этой работе хорошо систематизированы методы обработки и анализа электроэнцефалографических процессов. При этом автор предоставляет исследователю возможность выбора методов регистрации и обработки ЭЭГ в зависимости от задачи исследования. Эти методы достаточно широко используются и на практике.
Необходимо отметить, что во всех общепринятых и компьютеризированных методах анализа и даже при полиграфической регистрации сохраняется традиционный подход, который сводится к поиску характерных изменений аналоговой кривой, определению амплитудных и их интервальных показателей и статистической обработке. Компьютеризация обработки, по существу, является автоматизацией способов, «добытых вручную» не одним поколением электрофизиологов. Поэтому, отдавая должное систематизации известных методов обработки электрической активности, представленной в монографии А. П. Кулаичева, следует признать, что в этих методах анализа ЭЭГ современный системный подход и методы моделирования не используются. Вообще же работ по системному анализу ЭЭГ с использованием корреляционного анализа частот ЭЭГ немного19202122.
Последние годы в связи с развитием современных компьютерных средств наблюдается значительное увеличение числа публикаций по анализу ЭЭГ как с расширением диапазона частот, так и с исследованием неоднородности традиционно выделяемых ритмов. Накапливаются корреляты функциональной значимости расширенного диапазона частот с патологическими состояниями, перестройкой в деятельности мозга в разные стадии сна, при воздействии различных внешних факторов, в том числе фармакологических, при выполнении многих видов операторской работы и когнитивной деятельности. Обстоятельные сводки по этим публикациям даны в обзоре А. И. Федотчева и др. 23
Показано, что методы спектрального анализа могут лишь отражать парные связи между потенциалами при разных отведениях, методы картирования – представить визуальную картину спектров, но не пространственно-временную взаимосвязь процессов. Что касается корреляционных связей между частотами разных областей мозга, то собственный опыт использования корреляционного анализа состояния гемодинамики, показанный выше, убедил в их лабильности и наличии очень сложной картины так называемых откликов, которые не позволяют с сколько-нибудь надежно представить взаимосвязь процессов.
В последнее время интенсивно разрабатывается два методических направления анализа ЭЭГ:
- технологии разделения источников сигнала, в частности, метод независимых компонент24;
- статистические методы анализа большого числа переменных при небольшом числе наблюдений, включающие рандомизированные методы и технологию управления частотой ложных позитивов (Fals Discovery Rate)2526.
Однако ни один из существующих методов анализа ЭЭГ не является прямым методом системного анализа. Весь опыт электроэнцефалографии показывает невозможность проведения формальной границы между нормой и патологией. И все же в последние годы интенсификация разработок признаков состояния здоровья привела к формированию трех основных классов:
- нормальной ЭЭГ (ЭЭГ здорового человека);
- пограничной между нормой и патологией;
- патологической (анормальной).
При этом пограничная область просто включает слабо выраженные патогномоничные признаки. Например, появление одного из 4 видов эпилептиформной активности (спайков, острых волн, комплексов спайк-волна и др.). Ф. Я. Золотарев27 выделяет три основных паттерна ЭЭГ в норме по степени выраженности альфа-ритма:
- организованный, регулярный;
- нерегулярный;
- плоский, т. е. низкоамплитудный.
Е. А. Жирмунская28 полагает, что нормальная ЭЭГ можно быть определена по отсутствию межполушарной асимметрии в фоновой ЭЭГ, пароксизмальной активности и фокуса патологической активности. При этом Е. А. Жирмунская считает, что состоянии пассивного бодрствования у абсолютного большинства практически здоровых может быть 4 типа паттерна ЭЭГ:
- организованный, где основной компонент – альфа-ритм;
- гиперсинхронный (высокий индекс регулярных колебаний);
- десинхронный (резкое уменьшение количества альфа-волн;
- дезорганизованный (нерегулярная альфа-активность).
В основных типах автор выделяет еще и группы.
Однако, несмотря на обширную литературу по ЭЭГ-исследованиям в норме и различной патологии головного мозга, до сих пор нет четкого определения нормальной ЭЭГ и единого мнения о вариациях паттерна ЭЭГ практически здорового человека. В исследованиях функционального состояния центральной нервной системы большой интерес представляют исследования симметрии электрической активности полушарий головного мозга [Е. А. Жирмунская, 1989].
Асимметрия полушарий подробно рассмотрена в рамках функциональной асимметрии сенсорной, моторной и нервно-психической деятельности человека [Н. Н. Брагина, Т. А. Доброхотова, 1988]. Основным методом в этих исследованиях является сопоставление психофизических функциональных нарушений, возникающих при повреждении правого или левого полушарий мозга. Однако к топографии патологических изменений ЭЭГ, которой посвящено большое число работ, по сути, это не имеет отношения. Как указывалось выше, Е. А. Жирмунская полагает, что в норме межполушарная асимметрия ЭЭГ отсутствует. Функциональная асимметрия полушарий по существу сводится к пространственно-временной дезорганизации целостной деятельности человека и его психики. Установленное функциональное различие в пространственно-временной организации, составляющей наиболее фундаментальную характеристику целостной нервно-психической деятельности человека, до сих пор не нашло соответствующего отражения в электроэнцефалографических исследованиях.
В известной мере это определяется ограничениями существующих методов анализа ЭЭГ. Так, методы картирования представляют визуальную картину спектров, но не пространственно-временную взаимосвязь процессов. Последнее особенно важным представляется для характеристики здорового (или практически здорового) человека. В рекомендациях Минздрава РФ по автоматизированной оценки ЭЭГ для определения групп риска патологии центральной нервной системы (1989) за основу оценки принято словесное описание ЭЭГ и ответов на функциональные пробы с унификацией текста заключения и терминологии. Первичное описание электрической активности мозга и ответов на пробы проводится на основании визуальной оценки записей с помощью приведенной кодировочной таблицы (списка эталонных формулировок, принятых в клинической электроэнцефалографии). Метод унифицирует анализ ЭЭГ и тем самым повышает его эффективность.
Авторы на основании накопленного опыта регистрации ЭЭГ выделяют ранние стадии нарушения функционального состояния мозга, что представляется полезным для дальнейшего развития и улучшения использования электроэнцефалографии. Однако классификация типов или вернее видов ЭЭГ здоровых людей представляется недостаточно обоснованной. Чтобы пояснить это, следует остановиться на некоторых принципиальных подходах к классификации типов ЭЭГ здоровых людей и описанных в литературе некоторых паттернах ЭЭГ здоровых людей. Со времен классических работ Мэгуна, Моруцции, Джаспера (1949-1965), как уже указывалось, исследователи различают два противоположных состояния биоэлектрической активности мозга: синхронизации и десинхронизации. Тогда же было выдвинуто предположение, что эти процессы определяются в основном двумя системами – ретикулярной формацией и структурами таламуса. Затем последовала противоречивая история длительных исследований взаимоотношения этих структур, изложение которой не входит в нашу задачу, тем более, как оказалось, эти процессы могут возникать в самых разных структурах мозга и связаны с самыми разными факторами.
Н. Н. Данилова29 связывает паттерны ЭЭГ с существованием «резонансных волновых генераторов», активность которых может быть усилена действием вспышек на резонансной частоте и развитием оптимального функционального состояния. К выводу о существовании трех волновых генераторов с резонансными свойствами пришли также и другие авторы. Полагают, что существуют две самостоятельные системы, два волновых генератора, работающих на низкой и средней частоте альфа-ритма (7-8 Гц и 9-10 Гц). Низкий оказывает тормозное влияние на средний. Полагают, что механизм обусловлен структурными особенностями — наличием фазово-специфических нейронов со свойствами авторитмичности, т. е. нейронов-пейсмейкеров.
Найдено, что у низкореактивных испытуемых доминируют низкочастотные генераторы. На этой основе Н. Н. Даниловой предложена модель пейсмейкера альфа-активности. Выдвинуто положение, что баланс активности низко- и высокочастотного альфа ритма определяет уровень функционального состояния. Чем выше средняя частота ритма, тем эффективнее деятельность. Снижение функционального состояния – смена среднего альфа-активности на низкочастотный. Н. Н. Данилова делает вывод, что функциональное состояние человека определяется балансом активности низкочастотного и среднечастотного альфа-генераторов, различающихся своей локализацией в мозге и структурной организацией. К этим выводам Н. Н. Данилова пришла, используя метод ритмического светового раздражения. Тем самым утверждается положение о существовании специального механизма регуляции функционального состояния через систему многих волновых генераторов биоэлектрической активности мозга, различающихся резонансными частотами. Можно полагать, что гармонический состав ритмов является показателем функционального состояния. К этим выводам мы еще вернемся в свете проведенного системного анализа ЭЭГ.
Однако опыт ряда исследователей показал, что основу ритмического механизма регулирования функционального состояния составляет синхронное вовлечение возбудимости структур мозга, образующих единую функциональную констелляцию в процессе общей ритмической модуляции. Нейрофизиологические и нейрональные механизмы, лежащие в основе ритмов ЭЭГ, детально рассмотрены в ряде фундаментальных обзоров3031. Считается точно установленным, что ритмическая структура ЭЭГ человека отражает свойства таламо-кортикальных цепей, что паттерны ЭЭГ имеют топографическую локализацию в соответствии с организацией нервной системы, и частотная выраженность определяется взаимодействием специфических и неспецифических структур, осуществляющих саморегуляцию32. В настоящее время представляется очевидным, что на основе структуры ритмов ЭЭГ возможна физиологическое определение и интерпретация различных функциональных состояний центральной нервной системы, хотя природа явления синхронизации и десинхронизации ритмов ЭЭГ до сих пор остается во многом недостаточно ясной.
Имеются основания полагать, что структура биоритмов мозга отражает потенциальное и текущее функциональное состояние адаптации. Обнаружено, что статистическая структура ЭЭГ различна у лиц с высоким, средним и низким уровнем адаптивности33. Указанные авторы при построении вероятностно-ориентированных графов на основе статистических данных и корреляционного анализа основных ритмов ЭЭГ групп людей с разным уровнем адаптивности показали, что в первой и второй группах имеет место устойчивый альфа ритм с устойчивой связью с дельта-, тэта- и бэта-ритмами. В третьей группе наблюдается неустойчивость амплитудно-частотных параметров ЭЭГ и нестабильность корреляционной связи между ними. Следует подчеркнуть вероятностный характер корреляционных связей, их высокую лабильность и усложненность из-за наличия многочисленных так называемых откликов, что сильно снижает их диагностическую значимость.
Нестабильность ритмов с резким сдвигом в высокочастотную область у лиц с низкими потенциями к адаптации, наблюдали С. И. Сороко, С. С. Бекшаев, Ю. А. Сидоров34. При этом в клинической картине доминировали неврологические расстройства, вегетососудистые дистонии и другие виды дизрегуляций. В экспериментальных исследованиях35 на кошках было обнаружено при нарастании длительности облучения прогрессирующее нарушение взаимодействия нейронных популяций коры, гипоталамуса, таламуса, ретикулярной формации и гиппокампа, с электронно-микроскопическими нарушениями синаптических терминалей. Большой интерес представляют работы В. С. Русинова с соавторами36 о структуре связей электрических процессов в коре головного мозга человека и ее нарушении при измененных состояниях. На этой основе была предложена диагностика функциональных состояний мозга человека. Позднее было показано, что с показателями функционального состояния мозга связаны и психологические показатели уровня сознания37. Этими авторами продемонстрирована когерентность высокочастотного диапазона – бета- и гамма-ритмов. При этом авторы говорят о хорошо выраженных межполушарных связей этих ритмов.
При переходе в состояние дремоты («провал мыслей») показано снижение амплитуды высоких частот, ослабление корковых связей, вплоть до их межполушарного разобщения, которое отмечается уже с 20 Гц. Авторы делают вывод о существовании связи высокочастотного ритма ЭЭГ с когнитивными процессами и полагают, что функциональное значение дельта-ритма и медленного альфа-ритма, появляющихся наряду с высокочастотными ритмами, состоит в организации «внутренней концентрации внимания». Как уже отмечалось, этой связи мыслительных процессов с ЭЭГ в последнее время уделяется немало внимания, что подтверждается поиском новых методов анализа ЭЭГ38.
Системный анализ электроэнцефалограммы
Вопросы самоорганизации электрической активности мозга и значения симметрии электрической активности полушарий мозга для состояния здоровья диктуют необходимость минимизации количества неизвестных и максимального использования накопленного опыта в электрофизиологии для интерпретации получаемых данных. Поэтому мы в своих исследованиях воспользовались общепринятыми методами регистрации ЭЭГ и ограничились общепринятыми параметрами ритмов ЭЭГ того и другого полушария.
Использовалась упрощенная комбинация лобно-затылочного отведения слева и справа для регистрации наиболее изученных ритмов, представленных в табл. 6. На практике довольно часто используют различные упрощенные варианты системы «10-20%»39. Электроды в лобной области (пункты Fl, F2) сдвигаются несколько назад, так что становятся пригодны для отведения активности передней височной доли обоих полушарий. В затылочной области по затылочной линии располагают два электрода в позиции О1, О2.
Краткое описание структуры полипараметрической модели подсистемы электрофизиологических процессов головного мозга человека
Для системного анализа электроэнцефалограммы предложена полипараметрическая когнитивная модель, построенная в соответствии с общей концепцией функционирования интеллектуальных и образных систем на основе фрактальной геометрии процессов. В системе используется унифицированный набор подобных физических параметров электроэнцефалограммы в абсолютных значениях с естественным масштабированием (табл. 1)
Таблица 1. Тест-параметры (по рекомендации Международной федерации общества электроэнцефалографии и клинической нейрофизиологии40
| Ритмы | Частоты кол/с | Амплитуды, μV | ||||||
| Дельта (δ) | Тэта (θ) | Альфа (α) | Бета (β) | Дельта (δ) | Тэта (θ) | Альфа (α) | Бета (β) | |
| Минимальные | 0,5 | 4 | 8 | 14 | 20 | 25 | 20 | 5 |
| Максимальные | 3,5 | 7 | 13 | Выше 14 | 30 | 35 | 60 | 20 |
| Средние | 2,5 | 5 | 10 | 20 | 20 | 30 | 40 | 12 |
Для построения полипараметрической геометрической стандартной модели фрактала ЭЭГ (рис. 1) абсолютные величины параметров (в соответствии с рекомендацией Международной федерации общества
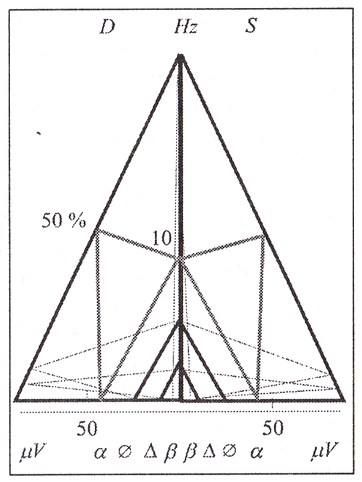
Для построения полипараметрической геометрической стандартной модели фрактала ЭЭГ (рис.1) абсолютные величины параметров (в соответствии с рекомендацией Международной федерации общества электроэнцефалографии и клинической нейрофизиологии) последовательно откладываются на катетах двух прямоугольных треугольников. Совокупность электрофизиологических параметров целого мозга замыкается в контуре равнобедренного треугольника при наложении катетов обоих прямоугольных треугольников, представляющих левое и правое полушария мозга. Катеты треугольников являются векторами временных (ось ординат) и амплитудных (ось абсцисс) параметров. Гипотенузы обоих треугольников представляют широко принятые в клинике индексы длительности каждого из ритмов ЭЭГ в процентах за 10-секундную эпоху регистрации для каждого полушария в отдельности.
В общей конструкции каждое полушарие имеет собственный интеллектуальный преобразователь системы, составляющий структурно-алгоритмический механизм функционирования системы, поскольку содержит всю основную характеристику ритмов (может содержать полный спектр частот) и позволяет осуществлять классификационные действия. Именно по этой конструкции возможно определение движения системы, т. е. структурно-функциональные изменения с оценкой величины энергии электрофизиологического процесса во времени в отдельных полушариях мозга. Интеллектуальный преобразователь осуществляет непрерывный анализ характера поведения абсолютных величин параметров и их соотношений и формирует в соответствии с этим необходимые классификационные действия (например, перевод данного пациента в систему клинических диагнозов).
Примечательно, что бета-ритм как бы не вписывается в строгий образ идеальной ЭЭГ. Действительно, по заключению известных специалистов, бета-ритм не является постоянным в нормальной ЭЭГ41.
Частота каждого ритма ЭЭГ в соотношении с его амплитудой в данной конструкции образует прямоугольный треугольник. Площадь такого треугольника имеет размерность кванта энергии (V*С) в соответствии с Международной системой единиц. По такому усредненному (за 10 с) кванту удобно сравнивать энергетику каждого из ритмов. По усредненным данным такие кванты ритмов составляют: для альфа-ритма 40μVС, тэта-ритма – 6μVС, дельта-ритма– 1μVC, бета-ритма 0,1μVС.
Для характеристики симметричных процессов наиболее удобной величиной может быть усредненный квант энергии ритма, умноженный на индекс ритма. Достаточно строгая характеристика процесса, наглядно представляемая такой конструкцией системы, дает основание для дальнейшего математического ее развития. В то же время она обладает веским достоинством наглядности.
При анализе соотношения временных и амплитудных параметров с использованием приемов проективной геометрии и общих принципов симметрии (ноу-хау метода) установлено, что минимальные, средние и максимальные величины параметров образуют пропорцию с коэффициентом, близким к 17=1,309 для левого и правого полушарий. Как показал наш опыт, с этим коэффициентом связана устойчивость модели системы и сохранность взаимоотношений ее элементов (параметров). Очевидно, что эти коэффициенты могут быть использованы в качестве критерия сохранности функционального порядка электроэнцефалографических процессов.
Для здорового человека без клинических признаков функциональных нарушений абсолютные значения находятся в пределах площади треугольника, т.е. могут варьировать в значительных пределах. Каково поведение соотношения основных ритмов в совокупности, предстояло выяснить. Длительный опыт электрокардиографических и реовазографических исследований показал, что изменения функционального состояния человека находят свое отражение в размере образа и (или) нарушении его контура относительно стандартной модели. Для диагностики состояния системы важна устойчивость соотношений ее параметров. При существенных отклонениях возникает напряжение, перенапряжение и сбой общей системы с соответствующими последствиями функциональных нарушений целого организма, т.е. возникновение разнообразной патологии.
Возможности практического использования предложенного способа анализа электроэнцефалограммы исследовали в сопоставлении с традиционными клинико-физиологическими методами анализа ЭЭГ.
При обследовании 12 работниц завода электронных приборов ЭЭГ регистрировали во фронтально-окципитальных стандартных отведениях потенциалов слева и справа; обработку сигналов ЭЭГ проводили общепринятыми методами с вычислением амплитудно-временных параметров.
Анализ результатов проводили методом клинической расшифровки ЭЭГ42 и на основе полипараметрической интеллектуальной системы с оценкой изменений соотношения параметров ЭЭГ. В качестве примера на рис. 2 приводим анализ ЭЭГ практически здорового человека с напряжением общего функционального состояния и анализ ЭЭГ больной с диагнозом дисфункции срединных структур мозга.

Как видно из рис. 2,a, т. е. в случае практически здорового человека, общий паттерн мало отличается от стандартного (модельного) паттерна ритмов, при этом коэффициент соотношения их составляет 1Г=1,24, т. е. отличается от «идеального» соотношения менее чем на 7%. В то время как на рис. 2,6 и паттерн значительно отличается от стандартного, и коэффициент соотношения ритмов составляет 1,03, т. е. его отклонение от инварианта более 20%, что хорошо соответствует клиническому диагнозу пациентки. Из 12 случаев наблюдения мы не обнаружили значимого расхождения между клиническим заключением и результатом системного анализа ЭЭГ. Хотя, конечно, это небольшое число для выводов о диагностической значимости каждого из новых признаков. Важным представляется наглядность картины, ее объективность и быстрота ее оценки, являющиеся значительным преимуществом метода полипараметрического когнитивного моделирования процессов.
Полученные результаты дают основания для более широкой апробации данного подхода к системному анализу ЭЭГ.
В табл. представлены результаты статистического анализа частоты основных ритмов электроэнцефалограммы студентов, проведенного старшим научным сотрудником нашей лаборатории В.А.Шаталовым. Энцефалограмму регистрировали при фронтально-окципитальном отведении в покое и коэффициенты соотношения основных ритмов определяли для левого и правого полушарий каждого студента (99 регистраций ЭЭГ выборки из 50 студентов 1-го курса).
Статистика фоновых ритмов электроэнцефалограммы и коэффициентов соотношения их (W) в правом и левом полушариях
Отведение =d
| Дельта | Тета | Альфа | Бета | W | ||
| N | Valid | 49 | 49 | 49 | 49 | 49 |
| Missing | 0 | 0 | 0 | 0 | 0 | |
| Mean | 1,510 | 5,396 | 10,824 | 16,447 | 1,2723 | |
| Std. Error of Mean | ,0608 | ,0678 | ,0912 | ,1619 | ,01209 | |
| Minimum | ,5 | 4,5 | 9,5 | 11,9 | 1,03 | |
| Maximum | 2,4 | 6,3 | 12,3 | 17,8 | 1,44 | |
| Percentiles | 25 | 1,300 | 5,000 | 10,350 | 16,450 | 1,2193 |
| 50 | 1,500 | 5,300 | 10,800 | 16,800 | 1,2677 | |
| 75 | 1,800 | 5,800 | 11,300 | 17,000 | 1,3381 | |
Отведение =I
| Дельта | Тета | Альфа | Бета | W | ||
| N | Valid | 50 | 50 | 50 | 50 | 50 |
| Missing | 0 | 0 | 0 | 0 | 0 | |
| Mean | 1,492 | 5,456 | 10,756 | 16,686 | 1,2991 | |
| Std. Error of Mean | ,0491 | ,0772 | ,0903 | ,0751 | ,01181 | |
| Minimum | ,5 | 4,3 | 9,3 | 14,8 | 1,15 | |
| Maximum | 2,3 | 6,8 | 12,0 | 17,8 | 1,53 | |
| Percentiles | 25 | 1,300 | 5,000 | 10,300 | 16,475 | 1,2419 |
| 50 | 1,500 | 5,500 | 10,800 | 16,800 | 1,2900 | |
| 75 | 1,800 | 5,850 | 11,300 | 17,000 | 1,3558 | |
Как видно из таблицы, частоты основных ритмов значительно различаются по абсолютным величинам (в 2-4 раза для медленных ритмов и на 20-25% для быстрых ритмов), в то время как коэффициенты соотношения ритмов не превышают 5% отклонения от инварианта, определяемого на модели. При этом различия коэффициентов для левого и правого полушарий не существенны.
Электрофизиологические процессы головного мозга обладают фрактальной самоорганизацией по принципу масштабно-инвариантного подобия и пропорциональность параметров является параметром порядка [А. И. Федотчев, 2003]. На основе представленной модели совокупность электрогенераторных структур целостного мозга правомерно представить как многоуровневую иерархическую систему (нейросеть) связанных нелинейных эндогенных осцилляторов – нейронов – с циклами разной периодичности в определенном (закодированном) порядке, эволюционно согласованным с экзогенными факторами. Такой подход может дать новые знания о нейронных процессах целостной системы головного мозга.
Развитие теории информации последних лет обратило внимание исследователей живых систем на кодовые соотношения элементов систем и связи между ними. Особый интерес для исследования закономерностей кодовых соотношений в различных системах представляет концепция компьютерной когнитивной графики (ККГ)4344. Несомненно, представленные кодовые соотношения параметров указывают на информационную основу функционального порядка как временную организацию сложных биохимических процессов.
Геометрическая структура модели фрактала дает возможность мероопределения соотношений, удовлетворяющие требованиям системного подхода и технологии извлечения знаний45. Следует отметить и высокую степень формализации получаемых результатов, что облегчает использование классификационных методов анализа. При этом геометрические образы легко интерпретируются на содержательном уровне и позволяют значительно снизить математические трудности при классификации.
Унифицированные схемы описания изображений открывают возможности для ее использования при интерпретации нейронных сетей различных структур мозга46.
Системный анализ электрофизиологических процессов на основе построенной модели показал широкие возможности такого инструментария. Модель обладает наглядностью иллюстративной графики и способностью генерации новых знаний о предмете: выявления информационных связей между элементами системы и закономерностей функциональной организации. Модель обладает дополнительными классификационными свойствами, способностью характеризовать качественно и количественно как объект в целом, так и его составные части.
Фрактальная геометрия импульсной активности нейрона
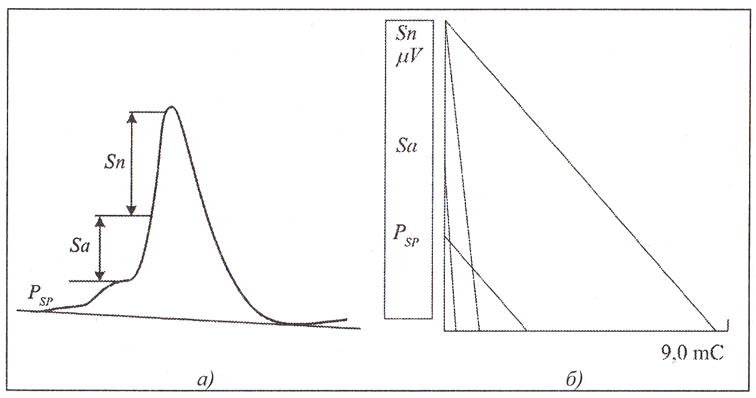
Представляется естественным попытаться посмотреть с позиции развиваемых представлений на электрическую активность отдельного нейрона. Для построения модели фрактала электрической активности единичного нейрона использовали наиболее репрезентативные из числа общепринятых временных и амплитудных параметров его импульсной активности: постсинаптический потенциал (Psp), спайк у основания аксона (Sa), спайк в теле нейрона (Sn)47. Такие представления об импульсной активности одиночного нейрона в известной мере являются стандартными, и поэтому речь идет отнюдь не об условном нейроне.
На рис. 3. представлены импульс нейрона и его модель. В прямоугольных координатах параметры импульсной активности нейрона даны в пропорциональном масштабе. Полученный треугольник, катетами которого являются временные (по оси абсцисс) и амплитудные (по оси ординат) параметры потенциала действия, является геометрическим фракталом импульсной активности нейрона. Анализ этой модели показал, что последовательное отношение отрезков длительности фаз активности свидетельствует об определенном временном порядке физиологического процесса возбуждения.
Эти результаты дают основание рассматривать проекцию временных параметров на прямой как метрику нейрональной активности. На основе общих принципов симметрии был определен инвариант соотношения всех параметров в их совокупности как наиболее устойчивое соотношение (И=1,309±0,113).
Площадь треугольника, катетами которого являются амплитуда потенциала действия и его длительность, можно рассматривать как квант электромагнитного потока, излучаемого нейроном при его работе.
Очевидно, можно только условно определить площадь треугольника модели как квант энергии или импульс силы (размерность последнего – ИС). Такая квантификация активности нейрона, по-видимому, была бы удобна для интегральной оценки его деятельности. Можно полагать, что характеристика электрических квантов нейрональной ткани и соотношение между ними могут явиться основным интегральным показателем функционального состояния нейронов, что может быть использовано как в экспериментальных условиях, так и при математическом анализе работы нейрональных популяций или сетей.
Электрофизиологические процессы головного мозга обладают фрактальной самоорганизацией по принципу масштабно-инвариантного подобия, и пропорциональность параметров является параметром порядка. Фрактальная геометрия моделей физиологических систем открывает возможности выявления кодовых отношений между параметрами физиологических функций и оценивать их взаимодействие количественно. На основе такой модели совокупность электрогенераторных структур целостного мозга правомерно представить как многоуровневую иерархическую систему связанных нелинейных эндогенных осцилляторов – нейронов – с циклами разной периодичности в определенном (закодированном) порядке, эволюционно согласованным с экзогенными факторами. Можно получить фрактальные визуализированные проекции сложной системы и строить многокомпонентную когнитивную модель.
Интерпретация полученных результатов на основе симметрийного подхода48 позволяет характеризовать обеспечение сохранности качества объекта в целом или его изменение, т. е. оптимальность и функциональную (закодированную) уравновешенность клеточно-тканевого объекта. Графическая модель потенциала действия дает наглядное представление о квантовой природе электрической активности нейронов как электрических осцилляторов с определенной частотой, вне предела которой такой квант в большей или меньшей степени деформируется.
Несомненно, это указывает на принципиальную основу функционального порядка как временную организацию сложных биохимических процессов «как особой корреляции, возникающей между биохимическими событиями и функциями клетки»49.
Можно высказать предположение, что на основе выявленного нами функционального порядка активности нейрона возникают макроскопические параметры функциональной организации мозга как нейрональной сети, имеющей тот же функциональный порядок.
Геометрическая структура модели системы дает возможность определять соотношения, удовлетворяющие требованиям системного подхода. Следует отметить и высокую степень формализации получаемых результатов, что облегчает использование классификационных методов анализа. При этом геометрические образы легко интерпретируются на содержательном уровне и позволяют значительно снизить математические трудности при классификации.
Унифицированные схемы описания изображений открывают возможности для использования соответствующего алгебраического подхода к их анализу50, который представляется перспективным для поддержки диагностических решений.
Воздержимся от более глубокой интерпретации полученных результатов, полагая, что для этого необходим более значительный объем экспериментальных данных.
Читайте также
- Системная электрофизиология
- Системный подход в биологии и физиологии
- Информационные процессы в живых организмах
- Методы системного анализа для моделирования живых систем
- Системный анализ электрокардиограммы
- Реоэнцефалография
Footnotes
- Жадин М. Н. Биофизические механизмы формирования электроэнцефалограммы. -М.: Наука, 1984.
- Lorente de No, R., Facilitation of mononeurons. – Amer. J. Physiology. 1953, vol. 113, p. 505.
- Elul, R., The Genesis of EEG // In: Intern. Rev. of Neurobiol, 1972, vol.15. New York and London, ed. С. C. Pfeiffer, J. R. Smithies.
- Джаспер Г. Риччи Г., Доун Б. Микроэлектродный анализ разрядов корковых клеток при выработке условных оборонительных рефлексов у обезьян // В кн.: Электроэнцефалографические исследования высшей нервной деятельности. – М. АН СССР, 1962, с. 129.
- Elul, R., The Genesis of EEG // In: Intern. Rev. of Neurobiol, 1972, vol.15. New York and London, ed. С. C. Pfeiffer, J. R. Smithies.
- Пурпура Д. Природа электрических потенциалов коры и синаптические структуры в коре головного мозга и мозжечка. // В кн.: Механизмы целого мозга. – М.: ИЛ, 1963, с. 9.
- Новикова Л. А. Современные представления о происхождении корковой ритмики и анализ электроэнцефалограммы // Сб.: Современные проблемы электроэнцефалографических исследований нервной системы. -М.: Медицина, 1964, с. 255-288.
- Elul, R., The Genesis of EEG // In: Intern. Rev. of Neurobiol, 1972, vol.15. New York and London, ed. С. C. Pfeiffer, J. R. Smithies.
- Elul, R., The Genesis of EEG // In: Intern. Rev. of Neurobiol, 1972, vol.15. New York and London, ed. С. C. Pfeiffer, J. R. Smithies.
- Жадин М. Н. Биофизические механизмы формирования электроэнцефалограммы. -М.: Наука, 1984.
- Гутман А. М. Биофизика внеклеточных полей мозга. – М.: Наука, 1980.
- Жадин М. Н. Биофизические механизмы формирования электроэнцефалограммы. -М.: Наука, 1984.
- Eccles, J. C„ The neurophysiological basis of mind. – Oxford, 1953.
- Винер Н. Кибернетика, или управление и связь в животном и машине. – М.: Мир, 1983.
- Жирмунская Е. А. Клиническая энцефалография. – М.: Мэйби, 1991.
- Davey, B. L„ Fright, W. R. Caroil, F. J. and Jones, R. D„ Express system approach to detection of Epileptiform Activity in the Medical and Biological Engineering, 1989,27,4, pp. 365-370.
- Жирмунская Е. А. Клиническая энцефалография. – М.: Мэйби, 1991.
- Кулаичев А. П. Статистическая диалоговая система. – М.: НПО «Информатика и компьютеры», 1991.
- Ливанов М. Н. Пространственная организация процессов головного мозга. – М.: Наука 1972.
- Суворов Н. Б., Василевский Н. Н., Медведева М. В. Нейроэффекты длительного действия микроволн: системное, нейрональное и электронно-микроскопическое исследование. – Радиобиология, 1987, т. 27, № 5, с. 674.
- Русинов В. С., Гриндель О. М. Оптимальная структура связей электрических процессов в коре большого мозга человека и ее нарушение при изменненых состояниях. -Журнал высшей нервной деятельности, 1988, т. 38, № 6, с. 985-1002.
- Василевский Н. Н„ Сидоров Ю. А., Суворов Н. Б. О роли биоритмологических процессов в механизмах адаптациии коррекции регуляторных дисфункций. — Физиология человека, 1993, т.19, № 1, с. 91-98.
- Федотчев А. И., Бондарь А. Т, Акоев И. Г. Ритмическая структура ЭЭГ человека: современное состояние и тенденции исследований. – Успехи физиологических наук, 2000, т. 31, №3, с. 39-53.
- Jung, Т.-P., Makeig, S„ Westerfield, M.. Townsend, J., Courchesne, E., Sejnov, Component analysis of singl-rial event-related potentials. Hum 2001,3, pp. 168-85.
- Wolpaw, J. R„ Bir, D. J., Pfurtscheller, G„ and Vaughan, T. M„ Brain-computer interfaces for communicate. – Neurophysiology, 2002 Jun, 113(6), pp. 767-791.
- Blunkertz, B., Vaughan, T. M., Schalk, G., Schlogl, A., et. al., The BCI Competition 2003: progress and perspectives in detection EEG single trials. – IEEE Trans Biomed ENG, 2004, Jun, 51(6), pp. 1044-1051.
- Золотарев Ф. Я. Значение параметров альфа активности электроэнцефалограммы для характеристики функционального состояния мозга человека // Автореф. дисс. … канд. биол. наук. – Л., 1979.
- Жирмунская Е. А. Клиническая энцефалография. – М.: Мэйби, 1991.
- Данилова Н. Н. Психофизиология. – М.: Аспект-пресс, 1998.
- Steriade, M. Gloor, P., Linas, R., et al. Basic mechanisms of cerebral thythme actievites // EEG clin.Neuruphysiology, 1990, vol. 76,4, pp. 481-508.
- Lope da Silva, F. H., Neural mechanism underlying brain waves: from neural membranes to networks//EEG Clin. Neurophysiology, 1991, vol. 79, no. 1, pp. 81-93.
- Sterman, M. B„ Physiological origins and functional correlates of EEG rhythmic activities: implications for self-regulation // Biofeedback and Self-Regulation, 1996, vol. 21, no. 1, pp. 3-33.
- Василевский Н. Н„ Сидоров Ю. А., Суворов Н. Б. О роли биоритмологических процессов в механизмах адаптациии коррекции регуляторных дисфункций. — Физиология человека, 1993, т.19, № 1, с. 91-98.
- Сороко С. И., Бекшаев С. С., Сидоров Ю. А. Основные типы механизмов саморегуляции мозга. – Л.: Наука, 1990.
- Суворов Н. Б., Василевский Н. Н., Медведева М. В. Нейроэффекты длительного действия микроволн: системное, нейрональное и электронно-микроскопическое исследование. – Радиобиология, 1987, т. 27, № 5, с. 674.
- Русинов В. С., Гриндель О. М„ Блдырева Г. И, Вакар Е. М., Майорчик В. Е. Диагностика и прогнозирование функциональных состояний мозга человека. – М.: Наука, 1988.
- Русалов В. М., Русалова М. Н, Митрофанов А. А. Психофизиологический подход к проблеме сознания. – Успехи физиологических наук, 2005, т. 36, № 4, с. 37-56.
- Асланов А. С., Гаврилова Н. А., Сологуб Е. Б., Хризман Т. Л. Пространственная синхронизация электрической активности мозга человека в норме и патологии. И Пространственная синхронизация головного мозга человека. – М.: Наука, 1973, с. 128-167.
- Осовец С. М., Гинзбург Д А., Гурфинкель В. С. и др. Электрическая активность мозга: механизм и интерпретация. – Успехи физиологических наук, 1983,т. 141,вып. 1,с. 103-150.
- Гусельников В. И. Электрофизиология головного мозга. – М.: Высшая школа, 1974.
- Жирмунская Е. А. Клиническая энцефалография. – М.: Мэйби, 1991.
- Жирмунская Е. А. Клиническая энцефалография. – М.: Мэйби, 1991.
- Поспелов Д. А. На пути к искусственному интеллекту. – М.: Наука, 1982.
- Зенкин А. А. Когнитивная компьютерная графика. – М.: Наука, 1991.
- Городецкий В. И., Самойлов В. В., Малов А. О. Современное состояние технологии извлечения знаний из баз и хранилищ данных. – Новости искусственного интеллекта, 2002, №4 (52), с. 3-9.
- Яхно В. Г., Нуйдель И. В., Иванов А. Е„ Беллюстин И. С. и др. Исследование динамических режимов нейроподобных систем. – Информационные технологии и вычислительные системы, 2004, № 1, с. 126-148.
- Катц Б. Нерв, мышца и синапс. – М.: Мир, 1969.
- Система. Гармония. Симметрия / Под ред. В. С. Тюхтина и Ю. А. Урманцева. – М.: Мысль, 1988.
- КарериДж. Порядок и беспорядок в структуре материи. — М.: Мир, 1985.
- Гуревич И. Б., Журавлев Ю. И., Сметанин Ю. Г. Построение алгебр изображений на основе дескриптивного подхода. Математические методы распознавания образов // Доклады IX Всероссийской конференции. – М., 1999, с. 33-36.