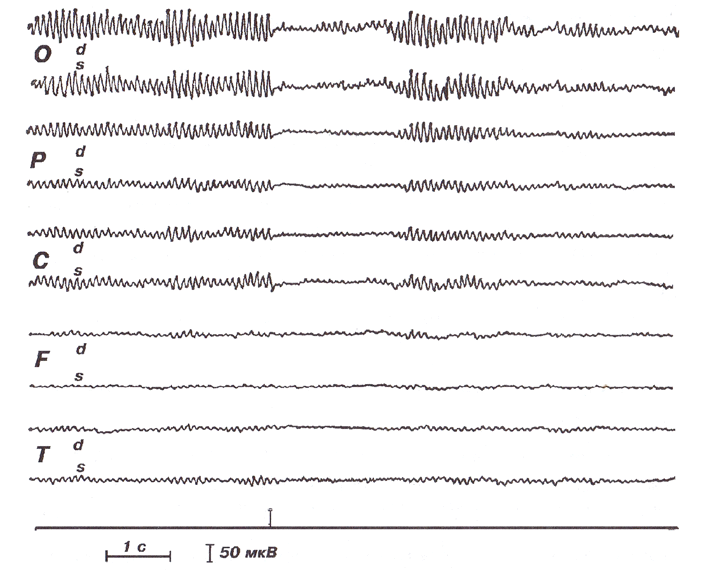
Регулярный a-ритм, модулированный в веретена, лучше всего выраженный в теменно-затылочных отделах. Реакция активации на вспышку света (отметка раздражения показана маркером).
Альфа-ритм или альфа-волны (англ. alpha rhythm, alpha wave, α-rhythm) — нормальная ритмическая активность ЭЭГ, регистрирующаяся в области первичных или вторичных зон сенсорной коры при открытых или закрытых глазах в состоянии покоя у 85-95% здоровых взрослых. При активации сенсорных областей коры альфа-ритм угнетается. У здорового человека альфа-ритм является доминирующим (т.е. преобладает по амплитуде) в области теменной, затылочной, а также сенсомоторной коры, где отдельно выделяют mu- или сенсомоторные ритмы. Частота альфа-ритма варьирует в пределах от 8 до 13 Гц. Синусоидальная форма амплитуды альфа-волн может варьировать, но у взрослых обычно около 30-50 мкВ. Блокируется или ослабляется при повышении внимания (в особенности зрительного) или мыслительной активности.
Таким образом, в современной литературе термин «альфа-ритм» применяется к любым компонентам ЭЭГ со следующими характеристиками:12345
- доминирующий ритм ЭЭГ в затылочно-теменной области с частотой 8-13 Гц;
- веретенообразность колебаний;
- снижение амплитуды при реакции зрительной и когнитивной активации
➥ Основная статья: Ритмы ЭЭГ
Функциональное значение альфа-ритма
Характеристики альфа-ритма — генерация в таламусе, теменно-затылочная топография, депрессия при открывании глаз — указывают на тесную связь альфа-ритма с функционированием зрительной системы. Однако физиологическая значимость альфа-ритма до конца не выяснена. Существует две полярные гипотезы. По мнению большинства исследователей, альфа-ритм – ритм «холостого хода» в зрительном анализаторе. Он возникает в отсутствие специфической зрительной афферентации и представляет собой ритм расслабленного бодрствования зрительной системы. Показано, что в условиях альфа-синхронизации реактивность коры значительно ниже, чем в фазу десинхронной ЭЭГ6. При наблюдении за данными кровотока также обнаружено, что в фазе синхронизации альфа-ритма потребление глюкозы значительно ниже, чем при десинхронизации. Эти данные позволяют считать, что реакция депрессии альфа-ритма отражает реакцию активации в зрительном анализаторе.
Согласно рассмотренной гипотезе аналогичная роль отводится ритмическим колебаниям в альфа-диапазоне, регистрируемым над проекционными зонами других анализаторов. Мю-ритм, подобно альфа-ритму, рассматривается как сенсомоторный «ритм холостого хода» — ритм покоя при отсутствии специфической афферентации в проприоцептивном анализаторе. Это предположение обосновывается распределением мю-ритма, депрессией на сенсорные стимулы или на пассивное движение. Тау-ритм височной коры, в свою очередь, отражает состояние функционального покоя в слуховом анализаторе.
Иное объяснение роли альфа-ритма было предложено Греем Уолтером (1966). Уолтер рассматривал генерацию альфа-ритма как отражение работы некоего сферического сканирующего механизма в зрительной системе: возрастание потенциала отражает период «очувствления» коры за счет восходящих таламокортикальных активирующих влияний на клетки коры.
➥ Подробнее: Таламус
ЭЭГ характеристики альфа-ритма
Лучше всего альфа-ритм выражен (доминирует) в затылочных и теменных отделах, по направлению кпереди амплитуда его постепенно уменьшается. Наибольшую амплитуду α-ритм имеет в состоянии спокойного расслабленного бодрствования, особенно при закрытых глазах в затемненном помещении. Для альфа-ритма характерна быстрота реакции, подавление альфа-ритма происходит при открывании глаз; при закрывании глаз амплитуда альфа-активности возвращается в норму, особенно в затылочной области.
Его амплитуда, хотя и является в среднем относительно постоянным параметром для данного индивидуума, весьма существенно колеблется во времени, как правило от 30 до 50 мкВ. Помимо изменений амплитуды, связанных с функциональным состоянием мозга, в большинстве случаев достаточно регулярно наблюдаются спонтанные изменения амплитуды, так называемые модуляции альфа-ритма, выражающиеся в чередующемся нарастании и снижении амплитуды волн с образованием характерных «веретен», длительность которых чаще всего колеблется от 2 до 8 с.
Модуляция – то есть правильное возрастание и убывание амплитуды альфа-ритма – выглядит на мониторе как горизонтальное веретено.
Регулярным считается альфа-ритм тогда, когда периоды (длины) волн отличаются не более чем на 0,5 Гц.
Количественной характеристикой является индекс альфа-активности. До 25% альфа-активность считается низкой, до 50% – средней, высокой – более 70%. Альфа активность считается нормальной, если она фиксируется в затылочно-теменной области с индексом не менее 60% или наблюдается во всех областях мозга с индексом не менее 50%.
По морфологическим свойствам альфа-ритм представляет собой синусоиду, но у молодых пациентов он может прерываться острыми отрицательными колебаниями, то усиливаться, то ослабляться, вызывая появление «эффекта биения». Подобный ритм может встречаться у одних людей и отсутствовать у других. Этот ритм называют «веретенообразным» (spindles). Он особенно заметен при исследовании сна у пациентов.
➥ Основная статья: Визуальные характеристики ЭЭГ
Факторы, влияющие на альфа-ритм
При повышении уровня функциональной активности мозга (напряженное внимание, интенсивная психическая работа, чувство страха, беспокойство) амплитуда α-ритма уменьшается, и часто он полностью исчезает. На ЭЭГ появляется высокочастотная нерегулярная активность. Автоматический анализ показывает, что в спектре мощности ЭЭГ исчезает доминантный пик в области a-ритма и спектр уплощается с равномерным распределением мощности по всем основным частотам, что подтверждает физиологическую интерпретацию этого феномена как десинхронизацию активности нейронов.
При кратковременном, внезапно возникающем на фоне покоя внешнем раздражении (особенно вспышке света) эта десинхронизация возникает резко, и в случае, если раздражение не носит эмоциогенного характера, наблюдается достаточно быстро (через 0,5-2 с) восстановление α-ритма (рис.). Такая реакция на ЭЭГ имеет несколько названий в зависимости от значения, которое в нее вкладывается: «реакция активации», «ориентировочная реакция», «реакция угасания α-ритма», «реакция десинхронизации». Реакция активации возникает как следствие появления нового фактора в окружающей обстановке, требующего дополнительной мобилизации активности организма и ориентации его в новой ситуации.
При повторных предъявлениях одного и того же стимула реакция активации постепенно слабеет и через некоторое время практически полностью угасает. Скорость угасания реакции зависит от субъективной значимости стимула, а также от внутреннего состояния организма, в связи с чем она может служить в определенных ситуациях мерой оценки состояния мозга. Ее наличие или отсутствие может быть дополнительным критерием степени реактивности мозга, показателем влияния нейротропных фармакологических средств. В частности, под влиянием нейролептика дроперидола наблюдается закономерное угасание реакции активации на редкие и нерегулярные световые стимулы, которые у человека в обычном состоянии вызывают реакцию активации, что может служить объективным критерием глубины нейролептического эффекта7.
Возраст. У большинства детей к трем годам альфа-ритм имеет частоту 8 Гц, у детей старше 10 лет – 10 Гц. У взрослых людей альфа-ритм становится доминирующей формой активности и преобладает во всех областях коры, средняя частота равняется 10–10,5 Гц8. Альфа-ритм (его частота и амплитуда) меняется с возрастом, что является отражением возрастных изменений в головном мозге человека при старении. С возрастом частота альфа-ритма замедляется, а у людей старше 80 лет альфа-ритм в среднем имеет частоту 7,5 Гц.
Половые различия. Альфа-ритм не имеет половых различий и одинаков у мужчин и женщин. Частота и амплитуда альфа-ритма зависят от фазы менструального цикла у женщин. Частота альфа-ритма увеличивается при уменьшении амплитуды в предменструальной фазе, а замедление частоты и увеличение амплитуды фиксируется во время месячных (Chang B., et. al., 2011 г.).
При повышении температуры тела увеличивается частота альфа-ритма. Онкологические заболевания, при которых наблюдается гипотермия, сопровождаются уменьшением частоты и амплитуды альфа-ритма.
Сердечный нервный узел (пейсмекер-водитель ритма) при его активации также повышает частоту альфа-ритма более чем на 1-2 Гц. Это происходит за счет усиления сердечного выброса, который приводит к повышению альфа-ритма.
Лекарственные вещества и лечение различными препаратами могут замедлять альфа-ритм.
Нарушения альфа-ритма и варианты нормы
Основная статья: Нарушения альфа-ритма: депрессия, асимметрия, дезорганизация, нерегулярность
⚠️ При отклонении от нормы характеристик (нерегулярность, дезорганизованность, нетипичное зональное распределение и др.) вместо альфа-ритма рекомендуется использовать более широкий термин альфа-активность.
В норме частота альфа-ритма у взрослых людей в затылочной области составляет от 8 до 13 Гц. Снижение частоты наблюдается при нарушении мозгового кровотока, т.е. уменьшении кровоснабжения в том или ином участке головного мозга. Частота ритма может временно изменяться в сторону увеличения при открытии глаз, переходя в ритм, похожий на бета-активность, который имеет частоту 13-30 Гц. Такой феномен называется «альфа-пиком» (alpha squeak).
Признаками патологии альфа-ритма являются: отличия периодов соседних волн на 1-2 Гц, модуляция имеет нечеткие и беспорядочные формы, волна альфа-ритма – не гладкая, а заостренная, зазубренная.
Чаще всего альфа-ритм – это доминирующая частота, то есть имеет максимальную амплитуду по сравнению с другими волнами. У 25% здоровых, нормальных людей альфа-ритм плохо визуализируется или может быть зафиксирован только периодически. Его амплитуда тоже может варьировать у разных людей и даже изменяться на протяжении жизни, независимо от наличия болезней головного мозга. Низкая амплитуда альфа-ритма отмечается менее чем у 10% людей. Альфа-ритм максимально визуализируется при исследовании затылочной области и перемещается в лобные отделы мозга при засыпании.
Асимметрию (то есть различие альфа-ритма между правым и левым полушарием) лучше оценивать при сравнении картины от двух задних электродных отведений (теменных и окципитальных) при биполярной записи, при использовании электроэнцефалографа экспертного уровня, дающего референсное качество. Высокая амплитуда характерна для правого полушария головного мозга (20-60 микровольт от пика до пика). При использовании Р4-О2 отведения, нормальная амплитуда составляет 15-45 микровольт.
Более высокая амплитуда характерна для замедленного альфа-ритма. Если асимметрия альфа-ритма между правым и левым полушарием достигает 50%, то необходимо искать патологический процесс в головном мозге.
Если вольтаж альфа-ритма в левом полушарии головного мозга больше чем на 35% по сравнению с правым полушарием, то также стоит искать патологический процесс в головном мозге.
Высокоамплитудный альфа-ритм в височных областях свидетельствует о наличии эпилептиформных изменений. Иногда такой ритм в височных областях головного мозга некоторые авторы называют «третий ритм», в отличие от «заднего альфа-ритма» и «мю-ритма» центральных отведений
Унилатеральное (одностороннее) изменение альфа-ритма свидетельствует об ослаблении ипсилатеральной (расположенной на той же стороне) патологии, включая заднюю область головы (Bancaunds phenomen).
Парадоксальная альфа-активность возникает тогда, когда она возрастает в передних отведениях, но явление сонливости отсутствует.
История открытия альфа-ритмов
Немецкий психолог Ганс Бергер (Gans Beiger) был первым, кто наблюдал альфа-ритмы, зарегистрированные от поверхности головы человека. Он сделал запись этого типа ритмов с поверхности головы субъектов, сидевших спокойно с закрытыми глазами. Ученый опубликовал свое открытие в 1929 году в статье Uber das Elektrenkephalogramm das Menschen — «Поверхностная электроэнцефалограмма человека» — и назвал эти электрические феномены «волнами первого порядка», или «а (алъфа)-волнами».
Бергер далее показал, что альфа-ритмы блокируются после открывания глаз или во время некоторых типов умственной активности, ведя к появлению «волн второго порядка», или «β (бета)-волн». Результаты Бергера были позже подтверждены несколькими другими исследователями, в частности Adrian and Matthews, которые ввели блестящую концепцию (которая остается верной до настоящего времени), что различные сенсорные области мозга имеют собственные альфа-ритмы (ритмы «холостого хода»), которые, по их предположениям, представляют состояние «отдыха», или «праздности» этой области мозга. Несмотря на то что со времен Бергера в электрофизиологии накопилась масса знаний относительно феноменологии альфа-ритмов, множество гипотез относительно механизмов и функционального значения этих ритмов не сходятся в общую теорию. Эта двусмысленность, вероятно, отражает разнородность альфа-ритмов.
Обычно альфа-ритмы представляют как ритмы нормальной человеческой ЭЭГ с частотами в диапазоне от 8 до 13 Гц. Определение альфа-ритмов как отдельной категории мозговой ритмики также предполагает особое пространственное распределение, мощный ответ на блокирование соответствующего сенсорного входа и особый механизм генерации. Существует не один, а несколько вариантов альфа-ритмов. Этот факт был известен с 1950-х (упоминается известным английским электрофизиологом Грэем Уолтером, однако достаточно описан он только недавно, когда стали обычной процедурой методы картирования мозга человека.
Мю-ритмы
➥ Основная статья: Мю-ритмы
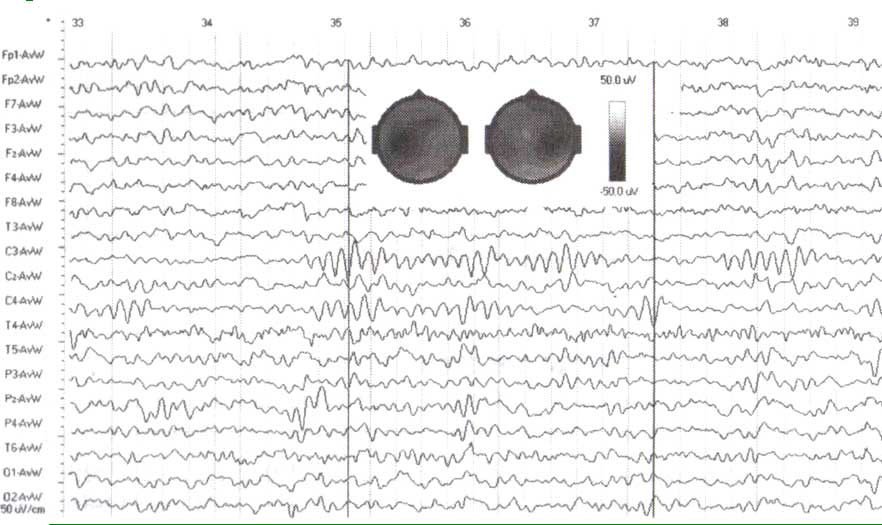
ЭЭГ, зарегистрированная у здорового взрослого субъекта в течение спокойного бодрствования с открытыми глазами. Обратите внимание на различные ритмы с частотой приблизительно 10 циклов в секунду в отведениях С3 и С4. Вставка — две топограммы потенциалов, снятых в два момента времени, указанных черными вертикальными линиями.
В литературе, посвященной нейробиообратной связи, была небольшая путаница при определении этого ритма. В некоторых статьях можно найти, что этот ритм определяется в диапазоне частот от 12 до 15 Гц. Это типичный пример несоответствующего применения к людям результатов, полученных в исследовании на животных. Дело в том, что сенсомоторный ритм интенсивно изучал в 1960-х годах Барри Стерман (Barry Sterman) из университета Калифорнии в Лос-Анджелесе (США). В экспериментах на кошках он обнаружил, что инструментально поощряемая генерация ритма ЭЭГ, регистрирующегося от сенсомоторной коры кошек, приводит к большей устойчивости к развитию приступов. Он обозначил этот ритм как сенсомоторный ритм. У кошек этот ритм находился в диапазоне 12—15 Гц. У людей, что недвусмысленно продемонстрировано исследованиями, частота сенсомоторного ритма изменяется в диапазоне 9—13 Гц. Барри Стерман сам указывал причину этой путаницы на различных конференциях и симпозиумах.
Затылочные альфа-ритмы
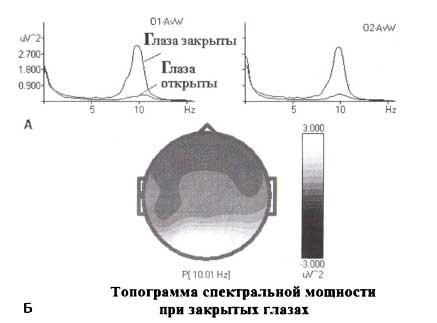
Высокоамплитудные ритмы, регистрирующиеся затылочными электродами от коры мозга человека, представляют другой пример альфа-ритмов. На рис. 3 изображен фрагмент ЭЭГ, зарегистрированной у здорового человека в состоянии с открытыми глазами. Во вставке на рис. 3 представлены спектры мощности для электродов в положениях О1 и О2. Как видно, затылочный ритм у этого человека проявляется в спектрах в форме острого пика. Визуально также можно оценить, что ритмы в О1 и О2-электродах этого человека синхронны. Синхронность — не обязательная особенность: у некоторых других здоровых людей ритмы в О1 и О2 могут быть независимыми друг от друга.
Затылочный ритм увеличивается (синхронизируется), когда человек закрывает глаза. Общие усредненные спектры в затылочных отведениях для группы здоровых взрослых в состояниях с открытыми и закрытыми глазами представлены на рис. 4. Как видим, закрывание глаз существенно увеличивает мощность ЭЭГ в диапазоне альфа-частот в затылочных областях. Эта синхронизация вызвана блокировкой зрительных входов к затылочным областям. Этот факт является одной из причин, почему затылочный альфа-ритм рассматривают как ритм «холостого хода» затылочной коры. Затылочный ритм обычно доминирует в записи ЭЭГ (также как и в спектрах) во время состояния спокойного бодрствования с закрытыми глазами. В некоторых редких случаях существуют не один, а два затылочных альфа-ритма. Они могут генерироваться в различных областях затылочной доли и могут иметь различные пиковые частоты.
Согласно литературе, частота затылочных ритмов изменяется с возрастом. На рис. 5 изображены спектры ЭЭГ, усредненные по группам здоровых людей в возрасте от 7 до 89 лет. Наблюдается небольшое увеличение средней частоты с 7 до 20 лет и небольшое уменьшение в последующие годы. Однако разница спектров ЭЭГ, представленная шириной пика, является большей, чем изменения с возрастом. Частота доминирующего альфа-ритма у здоровых пожилых людей приблизительно 10 Гц, так что частота затылочного альфа-ритма меньше чем 7,5 Гц обычно расценивается как умеренное отклонение.

Фрагмент электроэнцефалограммы, зарегистрированной у здорового взрослого субъекта в течение спокойного бодрствования с открытыми глазами. Обратите внимание на отчетливый ритм частотой приблизительно 10 циклов в секунду в отведениях О1 и О2. Вставка (вверху) - две топограммы потенциалов, снятых в два момента времени, указанных черными вертикальными линиями. Вставка (внизу) — спектры мощности ЭЭГ в отведениях О1 и О2.
Усредненные спектры для группы здоровых взрослых субъектов в состоянии спокойного бодрствования с закрытыми глазами по сравнению с состоянием с открытыми глазами. Внизу: топограмма разностного спектра для этих двух состояний.
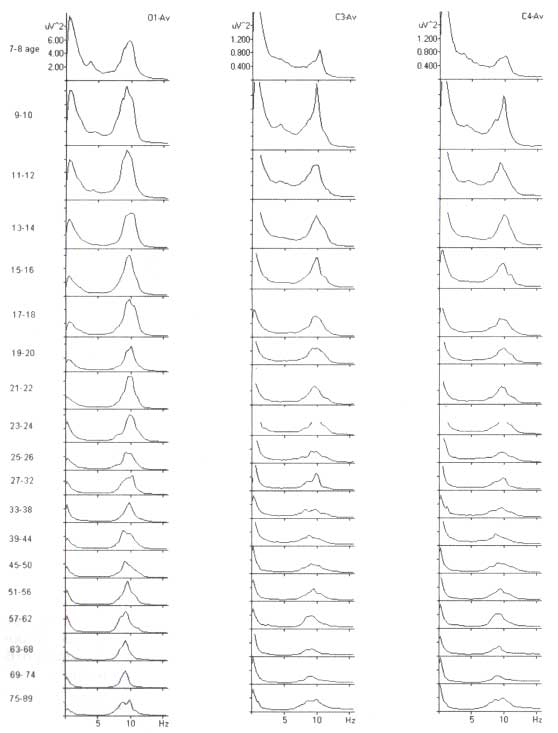
Обратите внимание, что усредненная мощность ЭЭГ в альфа-диапазоне заметно уменьшается с возрастом, в то время как частота максимума мощности усредненных спектров (частота ритмов) изменяется незначительно.
Теменной альфа-ритм
В некоторых редких случаях высокоамплитудные ритмы в диапазоне альфа-частот могут быть обнаружены в теменных областях с максимумом в Pz. Это рассматривается как нормальный вариант альфа-ритма. Этот ритм может увеличиваться в состоянии с закрытыми глазами, так же как затылочный альфа-ритм, хотя у некоторых людей мы наблюдали уменьшение теменного альфа-ритма в ответ на закрывание глаз. Теменной ритм может присутствовать в ЭЭГ вместе с затылочным ритмом и, по-видимому, независим от затылочного ритма (рис. 6). Частота теменного ритма обычно меньше частоты затылочного ритма, измеренная у того же человека. Суммарная мощность теменных ритмов возрастает с повышением трудности задачи и увеличивается в состоянии решения задачи по сравнению с состоянием спокойного бодрствования с открытыми глазами. Функциональное значение теменного ритма полностью не понято. Функциональный анализ сталкивается с двумя трудностями: 1) теменной альфа-ритм присутствует у немногих людей; 2) он маскируется соседним затылочным альфа-ритмом.
Недавний успех в развитии электромагнитной томографии открывает новые возможности трехмерной визуализации генераторов ЭЭГ-ритмов. Для этих целей успешно использовалась электромагнитная томография низкого разрешения (LORETA). В настоящее время LORETA становится обычным инструментом Мозгового картирования. Примеры двухмерных карт и соответствующих изображений LORETA различных типов альфа-ритмов (затылочного, сенсомоторного и теменного) у трех здоровых людей возраста 29, 19 и 18 лет представлены на рис. 7.
Три ритма, представленные на рис. 7, обнаружены у трех различных людей. Однако в некоторых случаях все три ритма могут быть найдены у одного нормального (здорового) человека. Более того, даже альфа-ритм с определенной локализацией (такой как затылочный) может иметь два и больше подритма с немного различной локализацией и с различными частотами.
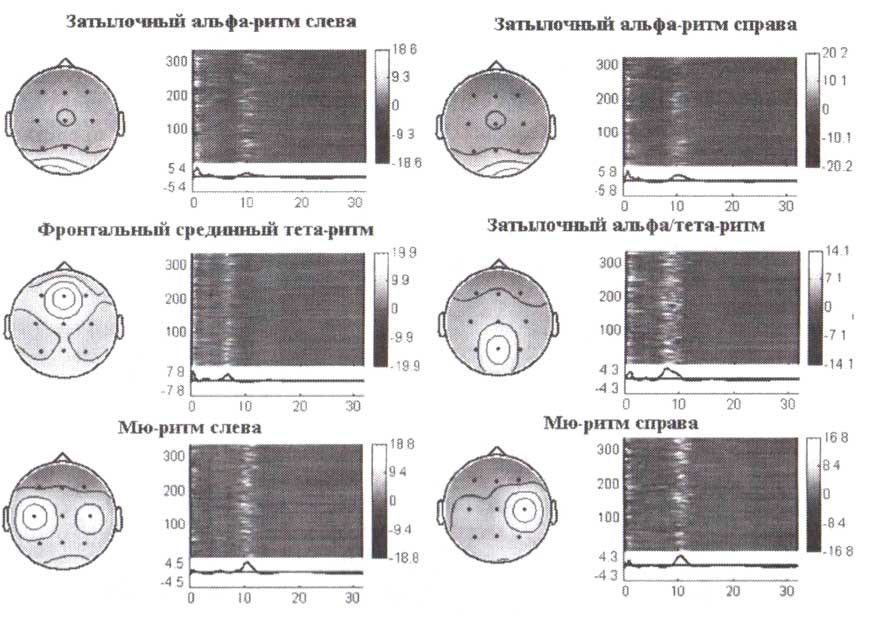
Анализ независимых компонент был применен к амплитудным спектрам ЭЭГ, вычисленным для более 300 здоровых субъектов, выполняющих зрительную двухстимульную GO/NOGO-задачу. Представлены шесть независимых компонент, соответствующих различным ритмам. Для каждой компоненты слева направо приведена топография компонент, представление соответствующей компоненты для отдельного субъекта, ниже — спектральные характеристики компонент.
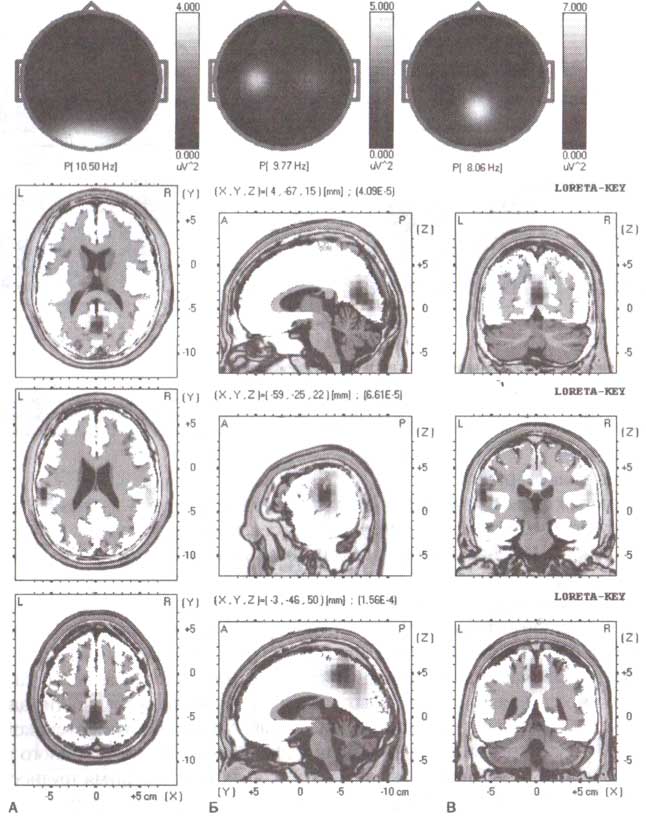
Примеры топограмм и соответствующих электромагнитных томографических изображений трех здоровых взрослых субъектов. Все субъекты находились в состоянии спокойного бодрствования с открытыми глазами. А. Индивидуум с затылочным альфа-ритмом. Б. Индивидуум с мю-ритмами. В. Индивидуум с теменным альфа-ритмом. Изображения LORETA отображают распределение генераторов ритмов в затылочно-теменной области, постцентральной сенсомоторной полосе и среднетеменной области соответственно.
Изменения в ответ на решение задач
Согласно теории генерации альфа-волн, представленной выше, любое увеличение сигнала на сенсорном входе должно подавить соответствующий альфа-ритм. Реакцию подавления называют реакцией десинхронизации, потому что высокоамплитудные альфа-волны можно рассматривать как результат синхронизации в нейронных сетях. Реакция десинхронизации альфа-ритмов в ответ на сенсорное возбуждение действительно наблюдалась во многих исследованиях. Эти исследования также внесли некоторую ясность относительно механизмов генерации альфа-ритмов и их функционального значения.
Вызванная десинхронизация мю-ритмов в ответ на двигательную активность

А. Вызванная десинхронизация, вычисленная для здорового взрослого субъекта, выполняющего двухстимульную GO/NOGO-задачу. Предъявляются пары изображений животных и растений в различных комбинациях, задача — нажать кнопку в ответ на два предъявленные подряд изображения животных. Обратите внимание на десинхронизацию мю-ритма с левой стороны, сопровождаемую вызванной синхронизацией в правой стороне во время подготовки к движению. Б. Вызванная десинхронизация, вычисленная для того же самого субъекта, выполняющего слуховую oddball-задачу. Значительная вызванная десинхронизация наблюдается при движениях в ответ на целевой стимул. Обратите внимание, что десинхронизация в одной области сопровождается противоположной реакцией — синхронизацией — в смежной области.
Ведущую роль в этих исследованиях сыграла группа исследователей из технического университета в Граце (Швейцария), возглавляемая Gert Pfurtscheller. В 1979 году он впервые опубликовал методологию параметрической оценки ЭЭГ-десинхронизации. Проанализированный параметр был назван вызванной десинхронизацией (ERD — event related desynchronization). В некоторых исследованиях вместо подавления альфа-активности при двигательных актах наблюдалось повышение мощности альфа-ритма. Эту реакцию назвали синхронизацией, а параметр, этому соответствующий, — вызванной синхронизацией (ERS).
Для пространственного картирования ERD/ERS обычно применяются монтаж локального среднего или монтажи подобные преобразованию Лапласа. Пример вычисления и картирования ERD показан на рис. 14, где представлены данные здорового взрослого человека. В спектрах ЭЭГ этого человека наблюдается отчетливый мю-ритм как в состоянии с открытыми глазами, так и в состоянии с закрытыми глазами. Этот испытуемый также выполнял двухстимульную GO/NOGO и слуховую oddball-задачи. Для оценки вызванной Десинхронизации в ответ на стимулы, предполагавшие двигательный ответ (нажатие на кнопку), анализ был проведен в соответствии с разработанным G. Pfurtscheller методом. В верхней части рис. 14 представлены вызванная десинхронизация/синхронизация между первым предупреждающим стимулом и вторым, который может быть или GO-стимулом (запускающим двигательную реакцию), или NOGO-стимулом (не требующим двигательного ответа). Можно заметить, что подготовительный период сопровождается десинхронизацией мю-ритма в контралатеральном полушарии и синхронизацией в ипсилатеральном полушарии. Внизу на рис. 14 представлены вызванная десинхронизация/синхронизация в ответ на слуховые стимулы: редкий стимул-мишень, предъявление которого требовало нажатия правым пальцем на кнопку, и стимул «не мишень», в ответ на который нажимать на кнопку не требовалось. Можно видеть, что предъявление только стимула-мишени вызывает сильную десинхронизацию мю-ритма.
Используя методологию вызванной десинхронизации/синхронизации в поведенческих исследованиях, связанных с сознательными движениями, группа из Граца сообщила о существовании нескольких подтипов роландических ритмов. Они показали, что мю-ритмы могут быть дифференцированы на основе их пространственной локализации. Например, области лица и ноги в сенсомоторной коре производят различные мю-ритмы. Группа Граца также показала, что мю-ритмы могут разделяться по частотам. Более низкочастотный (8—10 Гц) мю-ритм показывает широко распространенный неспецифический к типу движений паттерн вызванной десинхронизации, сходный для движений пальцев или ступней, тогда как более высокочастотный (10—13 Гц) мю-ритм демонстрирует более сфокусированный, специфический для типов движения паттерн вызванной десинхронизации, отличающийся для движений ступней и пальцев. Оба эти ритма блокируются перед и во время движений, и поэтому их можно рассматривать как мю-ритмы, хотя их реактивные компоненты различны.
Согласно данным, полученным группой из Граца, десинхронизация высокочастотного мю-ритма соматотопически специфична и топографически ограничена. Десинхронизация этого ритма начинается в области контралатеральной руки за две секунды перед началом сознательного движения ипсилатеральной руки. Десинхронизация часто сопровождается синхронизацией в центрально-теменных областях, и была названа «очаговой (локальной) десинхронизацией, окруженной зоной синхронизации». В целом этот термин описывает наблюдение, что десинхронизация ритмичной альфа-активности происходит не в изоляции, а может сопровождаться синхронизацией в соседних корковых областях. Области латеральной синхронизации могут соответствовать той же самой сенсорной модальности или другой модальности.
Чтобы объяснять явление «локальной десинхронизации, окруженной зоной синхронизации», G. Pfurtscheller предложил таламический селективный, воротный механизм. Он утверждал, что та же самая таламическая модель, которая использовалась для объяснения возникновения сонных веретен, может быть применена для объяснения генерации мю-ритма. Определяющую роль в его схеме играют взаимодействия между таламокорковыми клетками и тормозными нейронами таламического ретикулярного ядра. Последнее формирует топографически организованный тормозной механизм обратной связи, который способен управлять информационным потоком через таламус. Эту функцию управления таламических ретикулярных тормозных нейронов называют «селекцией, отбором,, или воротным механизмом». Когда ворота закрыты, таламические нейроны разряжаются в виде пачек спайков, которые проявляются в корковом мю-ритме. Когда ворота открыты, таламические нейроны разряжаются в тоновом режиме, который проявляется в десинхронизации мю-ритма. Согласно предложенному механизму, латеральное торможение, которое существует между ретикулярными нейронами, отвечает за явление «локальной десинхронизации, окруженной зоной синхронизации». Исходя из всего вышесказанного, можно заключить, что роландические мю-ритмы подавляются в ответ на реальные или воображаемые движения.
Вызванная десинхронизация затылочных ритмов в ответ на зрительные стимулы

Предстаалена усредненная вызванная десинхронизация, рассчитанная в диапазоне частот 8-12 Гц для релевантных и нерелевантных стимулов в двухстимульном GO/NOGO-тесте. Для примера была выбрана группа здоровых субъектов 12-13 лет. Обратите внимание, что релевантный стимул вызывает более длинный ответ, но с такой же амплитудой на начальных С1ндиях обработки, что и нерелевантный стимул. Обозначения: т1, т2—топограммы вызванной десинхронизации.
Похожие результаты показаны и для затылочных альфа-ритмов: они подавляются в ответ на зрительные стимулы. Кроме того, реакции в альфа-диапазоне отражают умственное напряжение, связанное с обработкой этих стимулов. Этот факт проиллюстрирован на рис. 15, где представлены данные, взятые из HBI базы данных. Вызванная десинхронизация/синхронизация для альфа-диапазона частот (8—12 Гц) вычислялась для группы здоровых людей в возрасте 12—13 лет. Они выполнили двухстимульное зрительное задание. Вызванная десинхронизация в затылочных областях в ответ на релевантные и нерелевантные стимулы представлена слева. Предъявление в паре стимулов первым релевантного стимула указывало на то, что испытуемые должны были ждать второго стимула (после которого требовался ответ GO или NOGO), в то время как предъявление нерелевантного стимула указывало, что вся проба должна игнорироваться. Как можно видеть, на первом этапе обработки информации (до 240 мс после стимула) оба типа стимулов выявляли одну и ту же десинхронизацию затылочной α-активности. Однако более поздние этапы обработки информации связаны с большей десинхронизацией затылочной альфа-активности для релевантного стимула по сравнению с нерелевантным.
Функциональное значение альфа-ритмов
Альфа-ритмы как ритмы «холостого хода» в ЭЭГ-активности
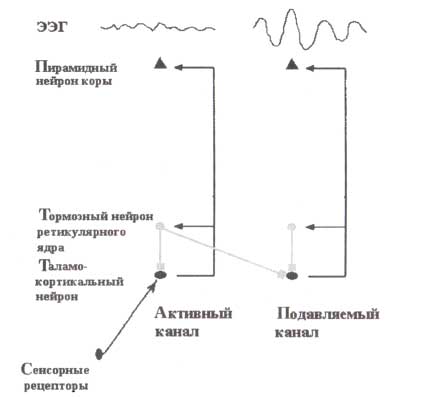
Схематично представлены два канала: активный (слева) и подавляемый (справа). Тормозные нейроны ретикулярного ядра в активном канале активируются и тормозят таламокортикальные нейроны в смежном канале. Нейроны в подавляемом канале гиперполяризованы и генерируют альфа-ритмы.
Как показывают фМРТ-исследования, величина синхронизации альфа-ритмов коррелирует отрицательно с метаболической активностью. Следовательно, альфа-ритмы рассматриваются как ритмы работы на «холостом ходу». Согласно этому представлению, мы могли бы ожидать, что появление альфа-ритмов в определенных сенсорных областях тормозит в них обработку сенсорной информации. Альфа-ритмы являются эпифеноменами, отражающими ритмические таламические разряды, передающиеся коре. Эти разряды отражают уровень нейронной деполяризации, которая, в свою очередь, отражает дисбаланс между возбуждающими и тормозными постсинаптическими потенциалами, приходящими в таламокоркальные нейроны (рис. 16). Если возбуждение достаточно сильно для того, чтобы деполяризовать таламо-кортикальные нейроны, то эти нейроны перестают генерировать пачки импульсов и переходят в другой режим, в котором информация от воспринимающих органов передается коре без всякого ограничения. Мы называем такой сенсорный путь активным каналом.
Латеральное торможение в активации альфа-ритмов
Если избыток торможения или недостаток возбуждения приводит мембранный потенциал в состояние гиперполяризации (то есть уровень возбудимости клетки падает), нейроны начинают разряжаться вспышками. Недостаток возбуждения может быть достигнут подавлением сенсорного входа на релейных (переключающих) ядрах таламуса, например, закрыванием глаз (для зрительной модальности). Избыток торможения может быть достигнут посредством латерального торможения, которое создается ретикулярными нейронами в близлежащем (активном) канале, возбуждающимися через связи от нейронов в активном канале как на корковом, так и на таламическом уровнях. Таламокортикальные нейроны в таком канале, который мы называем подавляемым каналом, производят разряды в частотах альфа-ритмов. Чтобы альфа-ритмы были зарегистрированы от скальпа, нейроны в подавляемом канале должны быть синхронизированы. При этом режиме синхронизации передача сенсорной информации к коре ограничена. Она полностью не блокируется как в течение ранних стадий сна, когда таламические нейроны генерируют сонные веретена, однако ограничена до некоторого уровня, такого что опознавание (обработка) стимула при этом режиме оказывается неполной.
И наконец, частота вспышек, произведенных в состоянии бодрствования в сетях подавляемого канала, определена уровнем поляризации таламокортикальных нейронов: чем больше они заторможены, тем медленней частота. В некоторых патологических случаях у взрослых эта частота может быть ниже 8 Гц.
Заключение
В состоянии спокойного расслабленного бодрствования с закрытыми глазами мозг человека продуцирует несколько типов ритмичной электрической активности в диапазоне альфа-частот (8—13 Гц). Обычно выделяют два основных типа альфа-ритмов, регистрируемых со скальпа: 1) затылочные альфа-ритмы, регистрирующиеся в затылочных или затылочно-теменных областях, и 2) роландический, или мю-ритм, регистрирующийся с сенсомоторной коры. Альфа-ритмы возникают в результате ритмичной активности таламических ядер: каждый ритм возникает в соответствующем таламическим ядре. Частота затылочного альфа-ритма немного изменяется с возрастом, достигая своего максимума в возрасте 15—20 лет.
Затылочные альфа-ритмы подавляются в ответ на зрительное возбуждение, в то время как роландические ритмы отвечают реакцией десинхронизации (уменьшения амплитуды) на реальные или воображаемые действия. Альфа-ритмы необходимо отделять и отличать от сонных веретен. Эти две различные категории ритмов наблюдаются в различных состояниях (во сне по сравнению с бодрствованием), имеют различное пространственное распределение (веретена сна имеют широкое центральное распределение, в то время как альфа-ритмы расположены около первичных сенсорных корковых областей), различные частоты (веретена сна — приблизительно 13—14 Гц в отличие от альфа-частот, которые изменяются между 8 и 13 Гц). Механизмы генерации сонных веретен подробно исследовались на модели животных. Что касается альфа-ритмов, механизмы их генерации известны не полностью. Мощность альфа-активности обратно пропорционально коррелирует с метаболической деятельностью в соответствующей корковой области, приводя к функциональному объяснению альфа-ритмов как нерабочих ритмов коры (ритмов расслабления и «холостого хода»).
Footnotes
- Ливанов М.Н. Избранные труды. Пространственно временная организация потенциалов и системная деятельность головного мозга. — М., 1989. — 400 с.
- Barry R..J., Clarke A.R.., Johnstone S.J. еt al. EEG differences between eyes-closed and eyes-open resting conditions // Clinical Neurophysiology. — 2007. — V. 118. — P. 2765-2773
- Klimesch W., SausengP., Hanslmayr S. EEG alpha oscillations: The inhibition-timing hypothesis// Brain Res. Rev. — 2007. — V. 53. – P. 63-88.
- Nunez P., Wingeier B., Silberstein R. Spatial-temporal structures of human alpha rhythms: theory, microcurrent sources, multiscale measurements, and global binding of networks // Hum. Brain Mapp. – 2001. – V. 13. – P. 125-164.
- Thatcher R.W., North D.M., Biver C.J. Intelligence and EEG phase reset: a two compartmental model of phase shift and lock// Neuroimage. — 2008. — V. 42, № 4. — P. 1639-1653.
- Гнездицкий В. В. Обратная задача ЭЭГ и клиническая электроэнцефалография. — М.: МЕДпресс-информ, 2004. — 624 с.
- Зенков Л.Р, Падалко В.В., Лавдовский В.С., Тонха Д.К. Нейрофизиологическое исследование премедикации предоперационного стресса дроперидолом. В кн.: Стресс и его патогенетические механизмы, «Штиинца», 1973, с. 74-76.
- Соколовская И.Э. Особенности ЭЭГ детского возраста / В кн. Нейрофизиологические исследования в клинике. – М.: Антидор, 2001. – С. 102-114.