Бета-ритм (Beta rhythm) — ритмическая ЭЭГ и МЭГ-активность более 13 Гц. Наиболее типичное определение: ритм в полосе частот от 13 Гц до 35 Гц, регистрируемый в лобно-центральных регионах головы в состоянии бодрствования. Иногда бета-ритм подразделяется на категории: низкий бета-ритм (бета-1) — от 13 до 21 Гц; высокий бета-ритм (бета-2) — от 21 до 30 Гц и гамма-ритм — активность с частотой более 30 Гц. Амплитуда лобно-центрального β-ритма варьирует, но обычно ниже 30 мкВ. Наблюдается подавление или ослабление бета-ритма при движениях или тактильной стимуляции контрлатеральной половины тела, что особенно явно видно на электрокортикограмме.
Лучше всего β-ритм регистрируется в области передних центральных извилин, однако распространяется и на задние центральные и лобные извилины. На ЭЭГ, отведенной от скальпа, в норме он весьма слабо выражен и в большинстве случаев имеет амплитуду 3-7 мкВ, т.е. всего в 2-3 раза превышает относительный уровень собственных шумов усилителя электроэнцефалографа, а при наличии артефактов ЭМГ может ими полностью маскироваться, β-ритм связан с соматическими, сенсорными и двигательными корковыми механизмами и дает реакцию на двигательную активацию или тактильную стимуляцию. При выполнении или даже умственном представлении движения β-ритм исчезает в зоне соответствующей корковой проекции.
Активность с частотой 40-70 Гц и амплитудой 5-7 мкВ иногда называют γ-ритмом. Поскольку столь быстрые колебания при весьма низкой амплитуде не могут быть достаточно эффективно воспроизведены электроэнцефалографами, использующими механические системы регистрации, этот ритм существенного клинического значения не имеет. Международная электроэнцефалографическая классификация рекомендует исключить понятие γ-ритма из употребления и использовать вместо этого выражение «высокочастотный β-ритм».
В последнее время рядом исследований было показано, что при использовании цифрового фильтрования и компьютерного вычисления спектральной мощности в диапазоне 35-45 Гц регистрируется особая, не зависимая от традиционного β-ритма, связанная с познавательными операциями, сензомоторной интеграцией, распознаванием тактильных стимулов и целенаправленными движениями «активность 40 Гц», оценка топического распределения которой и выраженность при соответствующих пробах дает ценную информацию о нормальных механизмах организации соответствующих высших психических функций и может служить диагностике их нарушений123.
Типы бета-ритмов
Бета-ритмы, проявляющиеся в отдельных пиках на спектрограммах, у здоровых людей обнаруживаются в различных областях коры. Бета-ритмы с частотой 13—30 Гц наиболее часто выявляются в лобных или центральных областях по сравнению с задними отделами коры. Могут быть выделены по крайней мере два различных бета-ритма: бета-ритмы с максимумами в спектрах ЭЭГ, расположенных в сенсомоторной зоне, — роландические бета-ритмы, и бета-ритмы, локализованные более фронтально, — лобные бета-ритмы. Они проявляются в форме пиков на индивидуальных спектрах. Однако только у немногих здоровых испытуемых выявляется отчетливый бета-ритм, и, как следствие, усредненные спектры в состояниях и с открытыми, и с закрытыми глазами имеют отчетливые максимумы в диапазоне бета-частот. Амплитуда бета-ритмов при измерении с объединенными ушными референтами меньше 20 мкВ. Асимметрия бета-ритмов обычно не превышает 35-40 %. Эмпирически любая асимметрия больше 50 % может считаться отклонением и заслуживает дополнительного внимания.
Роландические бета-ритмы
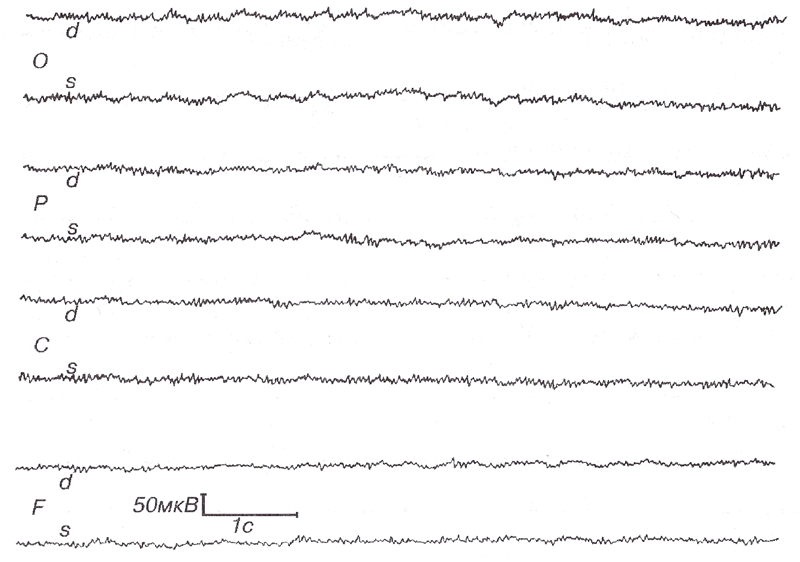
Роландические бета-ритмы наблюдаются как спонтанная активность в состояниях с открытыми и закрытыми глазами у здоровых испытуемых в сенсомоторных зонах (СЗ, Cz, С4). Хотя частота роландических бета-ритмов приблизительно 20 Гц (то есть в два раза чаще мю-ритма), их нельзя рассматривать как субгармонику низкочастотных колебаний мю-ритма. Действительно, как показали МЭГ-исследования, мю- и бета-ритмы имеют различные источники в первичной соматосенсорной и моторной коре соответственно (Хари и Salmelin, 1997, для обзора). Роландические бета-ритмы могут бьпъ обнаружены в ЭЭГ при отсутствии роландических мю-ритмов, а также вместе с роландическими мю-ритмами, но в различных отведениях (рис. 1). Частота роландического бета-ритма (максимум пика в спектрах) у разных людей может различаться, в широком диапазоне от 14 до 30 Гц. Кроме того, частота роландических бета-ритмов зависит от возраста.
Паттерны десинхронизации/синхронизации в ответ на двигательную активность
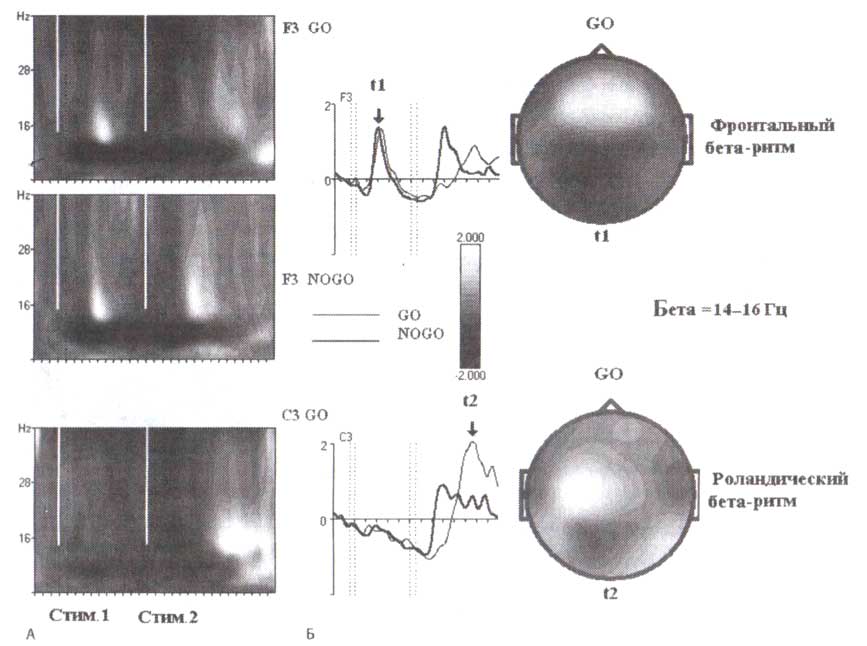
Роландический бета-ритм модулируется при выполнении различных двигательных и когнитивных задач. Пример динамики роландического бета-ритма в течение выполнения GO/NOGO-задачи в группе 15—16-летних подростков представлен на рис. 2. Как видно на рис. 2, роландический бета-ритм, зарегистрированный в СЗ, отчетливо подавляется во время нажатий пальцем после предъявления второго стимула в GO-пробах, так же как во время периодов подготовки, когда испытуемые готовились сделать движение, в пробах GO и NOGO, но не в игнорируемых пробах, когда испытуемому не надо было нажимать на кнопку. Обратите внимание, что уменьшение роландического бета-ритма (названное десинхронизацией) сопровождается уменьшением мощности ЭЭГ в альфа-диапазоне частот — десинхронизацией мю-ритма. За подавлением роландического бета-ритма, связанного с нажатием пальцем, следует обратная бета-синхронизация. Эта синхронизация имеет место во время продолжающейся десинхронизации мю-ритма, указывая на то, что роландические бета- и мю-ритмы имеют различную динамику. Необходимо подчеркнуть, что в исследованиях, представленных выше, предъявление стимулов служит точкой отсчета синхронизации. Однако так как роландические бета-ритмы связаны с движениями, разумно использовать как точки отсчета для вычисления вызванной десинхронизации сами движения, а не стимулы, вызывающие движения. В исследованиях, выполненных группой исследователей из Граца, возглавляемой Gert Pfurtscheller, испытуемых с множеством электродов, расположенных в зоне сенсомоторной коры, просили нажимать на кнопку произвольно и в удобном темпе. Исследования показали, что само движение сопровождается подавлением роландической бета-активности. Вслед за подавлением наблюдался сильный обратный эффект — бета-синхронизация.
Поскольку движение связано с глобальной активацией нейронов в сенсомоторной коре, мы можем предполагать, что роландическая бета-активность появляется, когда соответствующая нейронная система в сенсомоторной коре расслабляется после фазы сильной активации. Другими словами, увеличение роландической бета-активности — явление «отдачи», которое можно рассматривать как след постактивации.
Лобные бета-ритмы

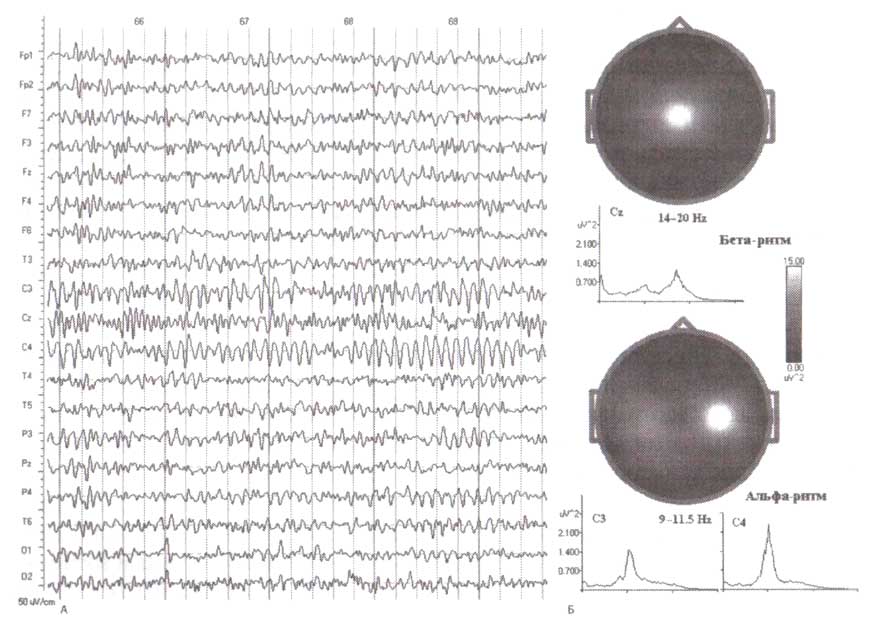
Второй тип бета-ритмов — лобные, которые обычно проявляются в спектрограммах лобных отведений как широкий пик с маленькой амплитудой. Такая форма спектра отражает низкую амплитуду этой ритмической активности и их нерегулярный паттерн. Бета-волны в монтаже с объединенным референтом редко, приблизительно у 2 % здорового населения, превышают 20 мкВ. Эти ритмы отличаются от альфа-ритмов центральных и задних отделов, которые обычно имеют более регулярные паттерны и выражаются в узком пике спектрограммы (рис. 3).
В F3-, Fz- и Р4-отведениях ЭЭГ спектры имеют отдельные максимумы в пределах 19 Гц.
Паттерны десинхронизации/синхронизации в ответ на когнитивные задачи
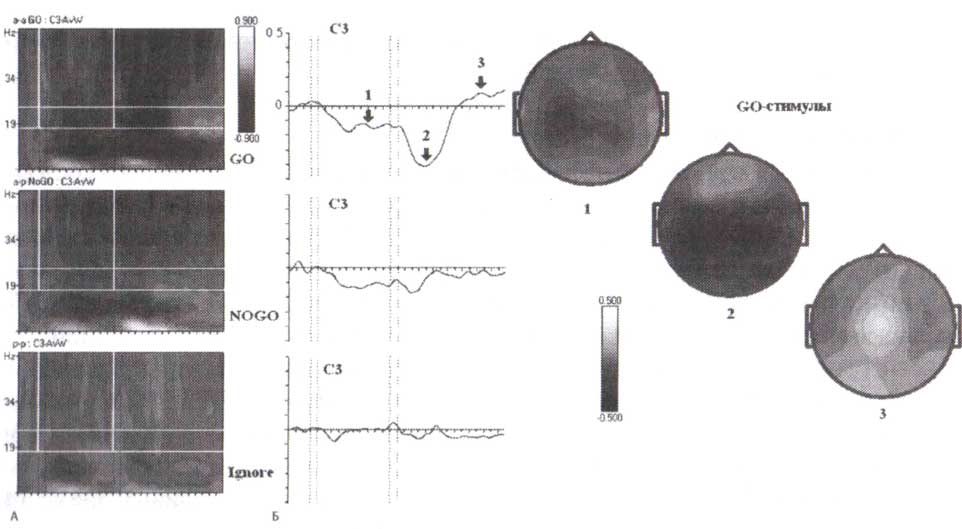
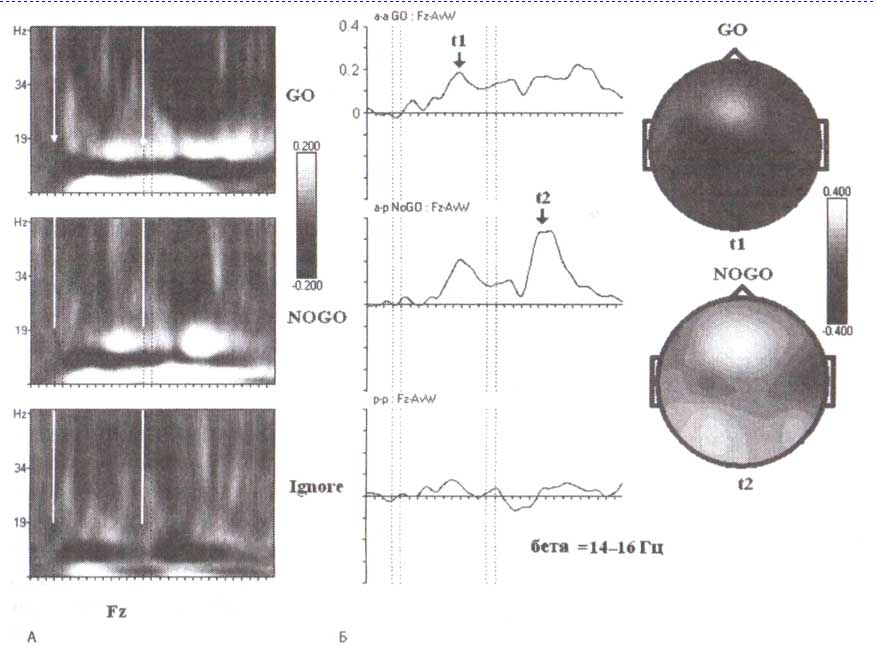
В отличие от роландических бета-ритмов (которые появляются при двигательных задачах) лобные появляются при решении когнитивных задач, связанных с оценкой стимула и принятием решения. У некоторых людей перед лобной бета-синхронизацией может возникать бета-десинхронизация. Но во всех случаях для развития лобной бета-активности требуется постстимульный интервал в несколько сотен миллисекунд. Лобный бета-ритм может присутствовать у человека вместе с роландическим (рис. 4). Первое предъявление стимула в GO- и NOGO-пробах вызывает начальную десинхронизацию с последующей синхронизацией отдачи в лобных областях (F3, F4). Напротив, бета-ритм в сенсомоторной зоне (СЗ) десинхронизируется во время подготовки к движению по отношению к синхронизации бета-ритма сразу после движения.
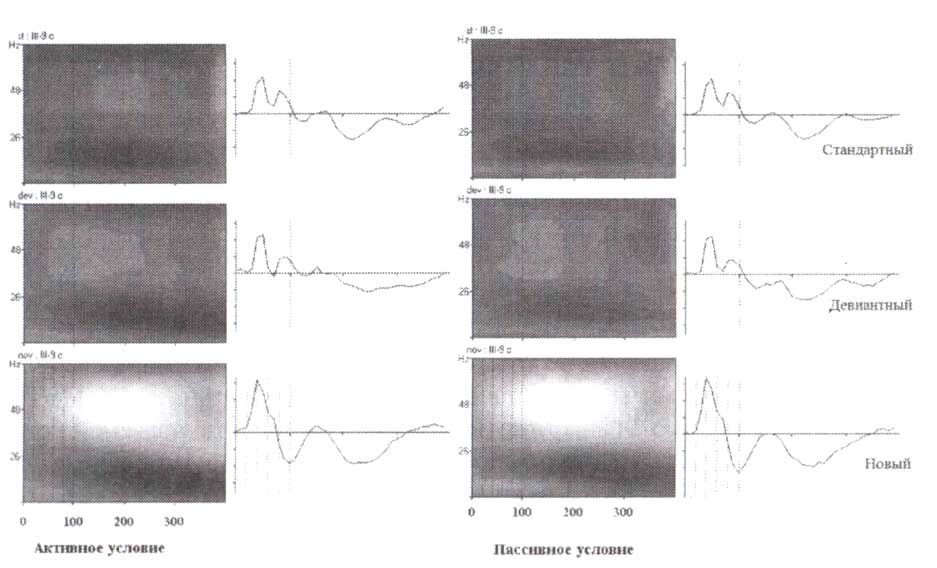
Мы выбрали для вышеупомянутого примера синхронизацию лобной бета-активности у одного здорового взрослого человека с отчетливым лобным бета-ритмом. Подобные данные могут быть получены на группах здоровых людей. На рис. 5, где представлены усредненные частотновременные реакции ЭЭГ-ответов при выполнении GO/NOGO-заданий группой 15—16-летних подростков, обратите внимание на то, что бета-синхронизация в ответ на первые стимулы в парах GO и NOGO начинается со значительной задержкой и достигает максимума в пределах 800 мс. Лобная бета-синхронизация не наблюдается в игнорируемых пробах (то есть в пробах, где предъявление первого стимула указывало на то, что вся проба должна игнорироваться).
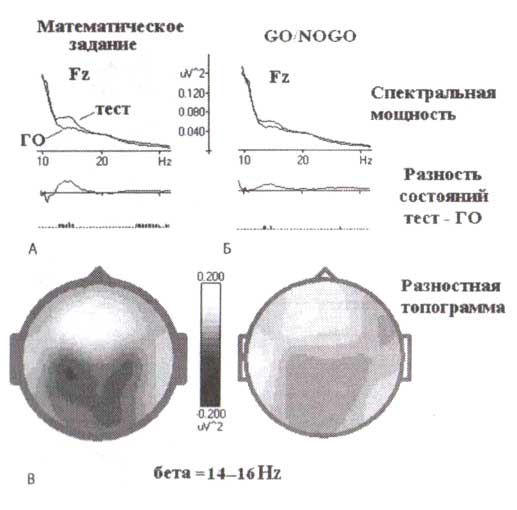
Синхронизация лобного бета-ритма, вызванная каким-либо заданием, может быть измерена как различие между суммарной мощностью ЭЭГ в бета-диапазоне при решении поставленной задачи и соответствующим параметром в состоянии спокойного бодрствования с открытыми глазами. Различия между спектрами в бета-диапазоне ЭЭГ при выполнении какой-либо когнитивной задачи могут являться мерой активации лобных долей при решении поставленных задач. Это отображено на рис. 6, где спектры ЭЭГ для двух различных задач (математической и GO/NOGO) наложены на спектры, вычисленные в состоянии спокойного бодрствования с открытыми глазами. Карта различий в бета-диапазоне частот показывает, что вызванная решением поставленной задачи бета-активность распределена в лобных зонах. Кроме того, степень увеличения лобного бета-ритма зависит от трудности задачи. Для сравнения изменения бета-активности, связанные с решением двух психологических задач, изображены на рис. 6. Первая задача — трудная математическая задача, которая требует интенсивного включения рабочей памяти. Вторая — более легкая GO/NOGO-задача. В ней волонтер должен реагировать только в 25 % проб во время выполнения простой задачи на различение. Трудность задач выражается в количестве ошибок, которые намного больше для математической задачи, чем для задачи GO/NOGO. Можно видеть, что трудная задача ведет к увеличению активности ЭЭГ в диапазоне частот от 14 до 20 Гц, вдвое больше, чем увеличение бета-активности при выполнении простой задачи.
Нейронные механизмы
Связь с корковой активацией
Существуют тесные отношения между, мощностью ЭЭГ в бета-диапазоне частот и метаболической деятельностью в соответствующей корковой области мозга человека. Это показано и для спонтанной ЭЭГ, и для вызванных ответов.
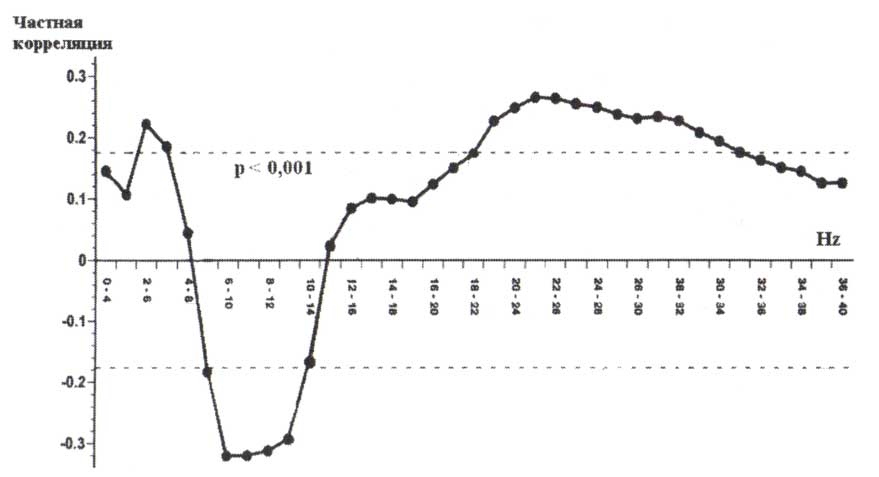
Одно из наиболее широкоизвестных исследований в этом направлении было сделано группой ученых из UCLA (Jan Cook и др., 1998). Они выполнили одновременную регистрацию многоканальной ЭЭГ и ПЭТ-исследование (с использованием радиоактивной воды) у здоровых взрослых людей в состоянии покоя и при выполнении простой двигательной задачи. Данные ЭЭГ были обработаны с использованием 3 различных монтажей, оценивались значения как относительной, так и абсолютной мощности ЭЭГ. Результаты исследования показали, что относительная мощность ЭЭГ более сильно коррелирует с перфузией, чем абсолютная мощность. Кроме того, вычисление мощности для пар биполярных электродов и усреднение мощности между электродными парами, разделяющими общий электрод привели к более сильной корреляции с перфузией, чем данные, вычисленные для монтажа с одним референтным электродом. На основе своих исследований авторы заключили, что: 1) топографическое картирование ЭЭГ может точно отразить локальное мозговую функцию способом, сопоставимым с другим методам; 2) выбор ЭЭГ-измерений и монтажа существенно влияет на степень, с которой ЭЭГ отражает локальную метаболическую активность. Результаты исследования продемонстрировали, что мощность ЭЭГ в бета-диапазоне частот, вычисленная с использованием локального среднего монтажа, положительно коррелирует с метаболической активностью в соответствующей локальной корковой области (рис. 7). Надо отметить, что мощность ЭЭГ в других диапазонах частот также коррелирует с метаболической активностью, но различными способами: альфа-активность, например, отрицательно коррелируете перфузией.
Тесные отношения между мощностью ЭЭГ и метаболической активностью мозга человека дают основание предполагать, что абсолютные значения изменений во всех диапазонах ЭЭГ могут служить индексом метаболической активации коры при решении определенных задач. Сравнение между fMRI и вызванной электромагнитной активностью было сделано в исследовании Singh (2002). Они сделали запись МЭГ- и fMRI-параметров при выполнении задачи на скорость распознавания букв при зашумлении сигнала. Изменения мощности МЭГ были нормализованы и усреднены между волонтерами. Результаты показывают, что вызванные решением поставленной задачи частотно-специфические изменения корковой синхронизации, обнаруженной на МЭГ, соответствуют тем областям мозга, в которых наблюдается вызванный корковый гемодинамический fMRI-ответ. Большинство этих изменений на МЭГ являлось вызванной десинхронизацией (ERDs) в бета- и альфа-диапазонах частот ЭЭГ.
Чувствительность к ГАМК-агонистам
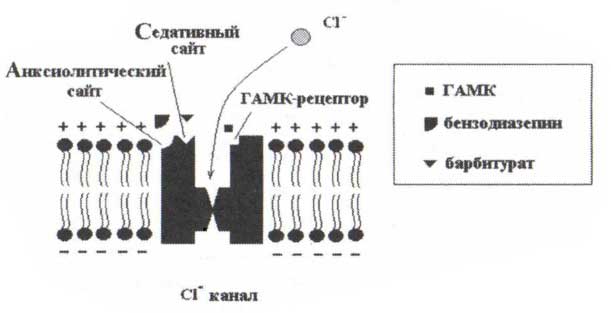
Самая поразительная особенность бета-ритмов — их чувствительность к ГАМК-агонистам. Действительно, мощность бета-активности увеличивается после приема барбитуратов, некоторых небарбитуратных успокоительных средств и легких транквилизаторов. Обратите внимание, что по контрасту с увеличением мощности бета-ритма средняя частота роландического бета-ритма после приема барбитуратов уменьшается (Jensen и др., 2005). На рис. 8 схематично объясняется действие вышеупомянутых средств. Показан СГ-лиганд-зависимый канал. Когда канал открыт, наблюдается инфузия (ток вовнутрь) в нейрон ионов СГ. Этот приток гиперполяризует мембрану и уменьшает вероятность возникновения нейронного разряда. Канал — сложная молекула, которая имеет несколько участков связывания: участок связывания ГАМК, зоны связывания седативных и анксиолитических веществ. Участок связывания ГАМК имеет сродство к молекуле самой ГАМК. Участок связывания седативных веществ имеет сродство к алкоголю или барбитуратам, таким как фенобарбитал. Участок связывания анксиолитических веществ связывается с бензодиазепинами, такими как валиум, либриум. Тормозные и возбуждающие синапсы отличаются по положению на мембране. Возбуждающие синапсы обычно располагаются на дендритах, в то время как тормозные синапсы расположены на теле клетки.
Идея о том, что колебания генерируются в результате коллективного поведения нейронов в сетях благодаря возвратному (циклическому) взаимодействию между ними, появилась давным-давно. Было предпринято много попыток смоделировать такие сети. Эти попытки находятся в рамках отдельной теоретической отрасли нейронаук, названной нейронными сетями.
Торможение в корковых цепях
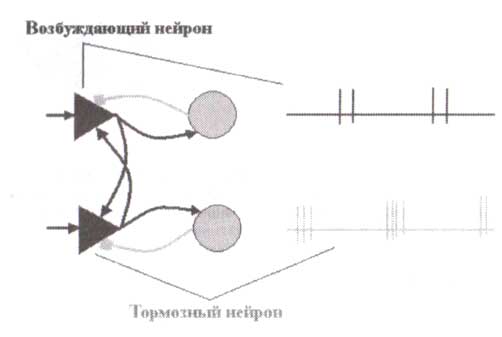
Исследование в смоделированных нейронных сетях показало, что тормозные нейроны могут играть определяющую роль в генерации колебаний. Общие сети, генерирующие колебания, состоят из двух типов реципрокно связанных нейронных цепей (сетей): тормозной и возбуждающей. Представьте теперь себе, что в такой сети заблокированы тормозные процессы. Благодаря рекуррентному возбуждению такая сеть будет лавинообразно самовозбуждаться по принципу положительной обратной связи. При такой обратной связи чем больше нейронов активно, тем больше они активируют другие нейроны через прямые связи, и тем больше они получают возбуждение через обратные связи. Положительная обратная связь может вести к сверхактивации сети. В действительности (в реальных условиях) такая сверхактивация ограничивается торможением. Тормозные нейроны оказывают регулирование действия на эти лавинообразные реакции: они периодически прерывают сверхактивацию сети. Частоты этих периодических прерываний сильно зависят от продолжительности тормозных постсинаптических потенциалов. Последовательность событий в такой нейронной сети схематично представлена на рис.9. Сеть содержит возбуждающие и тормозные клетки с взаимосвязями между ними и входами от внешних источников. Внешние входы служат источниками активации сети.
Выше приведена гипотетическая и упрощенная схема совокупных (групповых) нейронных осцилляций. Такое поведение нейронов возможно, если: 1) внешнее возбуждение достаточно сильно, чтобы вызвать лавинообразную реакцию; 2) возбуждающие связи достаточно сильны, чтобы удерживать возбуждение; 3) тормозные связи достаточно сильны для того, чтобы остановить лавину. Новое подтверждение причастности ГАМК-рецепторов в генерации бета-активности недавно было получено в совместных электроэнцефалографических/молекулярно-генетических исследованиях. Самая сильная связь с мощностью ЭЭГ в бета-частотах наблюдалась у короткого плеча хромосомы 4. Эта область содержит группу генов ГАМК А-рецептора (Porjesz и др., 2005).
Функциональное значение
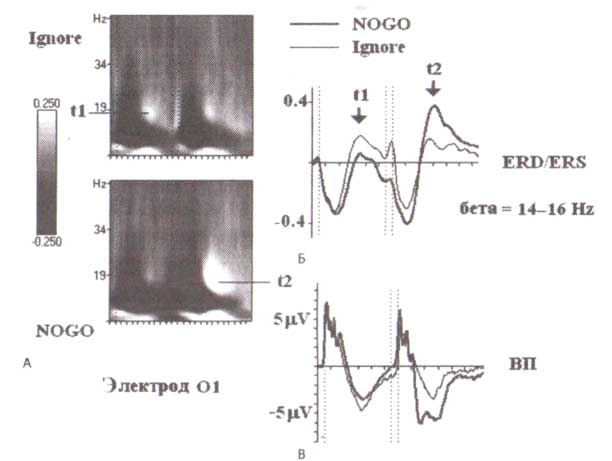
Бета-синхронизация в задачах, связанных со зрительной модальностью, может наблюдаться не только в соматосенсорных и лобных, но также и в затылочных областях коры (рис. 10). В этих областях бета-синхронизация происходит в ответ на игнорируемые или NOGO-стимулы, то есть на стимулы, которые сопровождаются «расслаблением» субъекта, но не на стимулы (типа GO-стимулы), которые нуждаются в дальнейшей обработке или реакциях. На рис. 11 показаны усредненные значения для wavelet-преобразования и вызванной де/синхронизации (ERD/ERS) в диапазоне бета-частот для левой затылочной области (электрод О1). Как видно на рисунке, бета-синхронизация в затылочной области сопровождается предшествующей ей значительной десинхронизацией и в альфа-, и в бета-диапазонах частот.
Бета-ритмы как следы постактивации
На рис. 11 самым важным фактом в отношении нейронных механизмов является наблюдение, что перед синхронизацией в бета-диапазоне наблюдалось увеличение отрицательных колебаний в вызванных потенциалах, зарегистрированных одновременно от того же самого электрода. Различия в негативных компонентах ВП в ответ на вторые стимулы между NOGO и игнорируемыми условиями возникают за 200 мс перед любыми различиями в бета-синхронизации. Это наблюдение указывает на то, что бета-синхронизация следует за изменениями в ВП со значимой задержкой. Положительные компоненты ВП (по крайней мере в затылочных областях в ответ на визуальные стимулы и для определенного возраста) связаны с возбуждающими постсинаптическими потенциалами, в то время как отрицательные компоненты ВП, по крайней мере частично, связаны с тормозными постсинаптическими потенциалами. Принимая это во внимание, мы можем заключить, что бета-синхронизация — след постактивации, которая следует за сильной активацией затылочной коры. Этот след постактивации возникает в результате включения тормозных взаимодействий в корковой нейронной сети (рис. 12).
Это заключение соответствует ряду наблюдений о синхронизации роландической и фронтальной бета-активности. Вспомним, что мы связали вызванную бета-синхронизацию с постактивационными (возвратными), эффектами в соматосенсорной и в лобной коре. Мы допускаем, что различные типы бета-синхронизации (в полосе от 14 до 30 Гц), возникающие при решении задач, в ответ на стимул или движение можно рассматривать как единичный феномен — след постактивации.
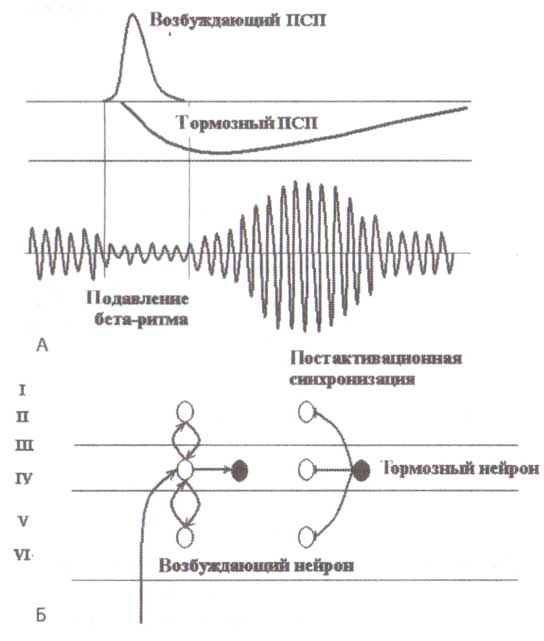
Перезагрузка обработки информации
Функциональное значение этого явления не ясно. Это может быть след памяти, который сохраняется в нервной системе, после ее активации. Однако согласно такому предположению, след должен быть сильнее для релевантных стимулов по сравнению с нерелевантными. Но мы наблюдаем прямо противоположное явление — след намного сильнее для нерелевантных стимулов, когда больше нет необходимости в использовании памяти. Самое вероятное объяснение наблюдаемых эффектов: бета-синхронизация как отдельное явление представляет собой процесс перезагрузки. Этот процесс необходим, чтобы стереть результаты предыдущей активации и соответствующих «вычислений» и подготовить систему к новым действиям.
Так или иначе бета-активность нужно рассматривать как отсроченный индекс корковой активации. Это происходит как реакция на сильную активацию корковых нейронных сетей, когда тормозные нейроны получают достаточное возбуждение от внешних входов и рядом расположенных возбуждающих нейронов. В течение этой стадии обработки сигнала тормозные нейроны начинают давать разряды, чтобы подавить сильную активацию.
Это торможение происходит циклично, и каждый цикл — цикл бета-активности, зарегистрированной со скальпа. В здоровом мозгу бета-активность может проявляться как «акт перезагрузки», который «стирает» последствия сильной активации в нейронных сетях, и это позволяет сетям обрабатывать информацию снова и снова.
При отклонении от нормы, когда это происходит почти постоянно и отражается в слишком высоком уровне бета-активности, она может рассматриваться как показатель гиперактивации и раздражения соответствующей корковой области. Синхронизация ритма в бета-диапазоне ЭЭГ также может увеличиваться, если облегчаются (потенциируются) тормозные взаимодействия, например, после употребления седативных лекарств и антидепрессантов.
Аномальные бета-ритмы
Потребность в нормативных базах данных
Как мы уже упоминали, генерация бета-ритмов связана с некоторыми специфическими состояниями коры. Эти состояния характеризуются высоким уровнем внешнего возбуждающего входа и относительно сильным внутрикорковым торможением. Эти состояния возникают во время постактивационной обработки информации в ответ на короткое предъявление стимула, когда тормозные нейроны включаются в работу, чтобы подавить деятельность нейронов, активных на первой стадии сенсорной обработки информации. Чтобы понять, является ли бета-ритм у отдельного человека слишком большим или слишком низким, необходимо сравнение данных по этому человеку с общей нормативной базой.
Когда у человека обнаруживается избыток или, наоборот, недостаток бета-активности, этот факт необходимо интерпретировать с осторожностью. Во-первых, для начала нужно оценить распределение бета-активности по поверхности головы. Нетрадиционному распределению бета-ритма, например в теменных или височных отведениях, должно быть уделено больше внимания, чем его распределению в сенсомоторных или лобных отведениях. Во-вторых, асимметрию бета-активности (выше 50 %) нужно рассматривать как дополнительный признак отклонения от нормы. В-третьих, реактивность наблюдаемого чрезмерного бета-ритма — важный показатель его функционирования. Если чрезмерная бета-активность у отдельного человека не синхронизируется в ответ на условие какого-либо задания так же, как в нормативной группе, то это наблюдение служит дополнительным индикатором отклонения от нормы. В-четвертых, необходимо принимать во внимание жалобы пациента, для чего необходимо вести протокол проводимой с ним текущей терапии и медикаментозного лечения.
Корковая ирритация (раздражение)
Увеличение бета-активности и соответствующей сверхактивации может происходить в областях, связанных с фокусом эпилептической активности, например во время предэпилептических аур. Другой пример: корковая гиперактивация может возникать при галлюцинациях. Слуховые галлюцинации — один из позитивных симптомов, связанных с шизофренией. ПЭТ-исследования показывают, что галлюцинации связаны с гиперметаболизмом в слуховых корковых областях. В соответствии с исследованиями, связывающими бета-активность с изменениями локального мозгового метаболизма, в МЭГ-исследованиях также было получено увеличение активности в бета-диапазоне ЭЭГ (12,5-30 Гц) в левой слуховой коре, сопровождавшее галлюцинации (Ropohl и др., 2003).
Иногда увеличение бета-акгивности может визуализироваться как веретенообразный бета-ритм. Этот паттерн, как известно, связан с «корковым раздражением», вирусными или лекарственными энцефалопатиями и эпилепсией. Такая неправильная бета-активность выглядит как увеличивающиеся и уменьшающиеся веретена. Избыток бета-акгивности наблюдается меньше чем у 10 % населения, страдающего синдромом нарушения внимания и гиперактивности (СНВГ).
Заключение
Полоса бета-ритма — это частоты ЭЭГ выше 13 Гц. Первым бета-ритм описал Ганс Бергер, эту мозговую электрическую активность связывали со сфокусированным вниманием. В полосе бета-ритма ЭЭГ можно выделить несколько его типов. Гетерогенность бета-ритма, разнородность подразумевает его традиционное деление на следующие поддиапазоны: низкочастотный бета-ритм — от 13 до 20 Гц, высокочастотный бета-ритм — от 21 до 30 Гц, гамма-активность — от 31 Гц и выше. Иногда выделяется отдельно особый тип «40 Гц активность». Этот тип бета-активности привлек большой интерес ученых в 1980-х. Исследования были сосредоточены на проблеме связанности восприятия. Было показано (и теоретически, и экспериментально), что сети тормозных промежуточных нейронов вовлечены в генерацию бета-ритмов. Причастность тормозных нейронов к генерации бета-ритма поддерживается идеей о чувствительности бета-ритмов к ГАМКергическим агонистам — фармацевтическим препаратам, которые подражают (имитируют) действию ГАМК, — основного тормозного медиатора в центральной нервной системе. ГАМК-агонисты, такие, например, как барбитураты и бензодиазепины, увеличивают мощность высокочастотных ритмов. В здоровом мозгу было показано, что бета-активность положительно коррелирует с метаболической активностью в корковой зоне, находящейся под записывающим электродом.
Footnotes
- Loring D.W., Sheer D.E., Largen J.W. Forty Hertz EEG activity in dementia of the Alzheimer type and Multi-infarct demenia / Psychophysiology, 1985, v. 22, №1, p. 116-121.
- Pfurtscheller G., Neuper Ch. Simultaneous EEG 10 Hz desynchronization and 40 Hz synchronization during finger movements/NeuroReport, 1992, v. 3, p. 1057—1060.
- Desmedt J.D., Tomberg C. Transient phase-locking of 40 Hz oscillation in prefrontal and parietal human cortax reflects the process of conscious somatic perception / Neurocsience Letters, 1994, v. 168, p. 126—129.