Мю-ритм (Mu rhythm) – ритм в полосе частот 8-13 Гц, представляющий собой аркообразные волны, регистрируемые в центральных и центрально-височных регионах скальпа в состоянии бодрствования. Амплитуда различна, но обычно ниже 50 мкВ. Наиболее явно блокируется или ослабляется контрлатеральными движениями, мысленным представлением движений, состоянием готовности к движению или тактильной стимуляцией. Обозначается греческой буквой µ. Синонимы: аркоидные (arceau), аркообразные (wicket), гребенчатые ритмы (применение не рекомендуется).
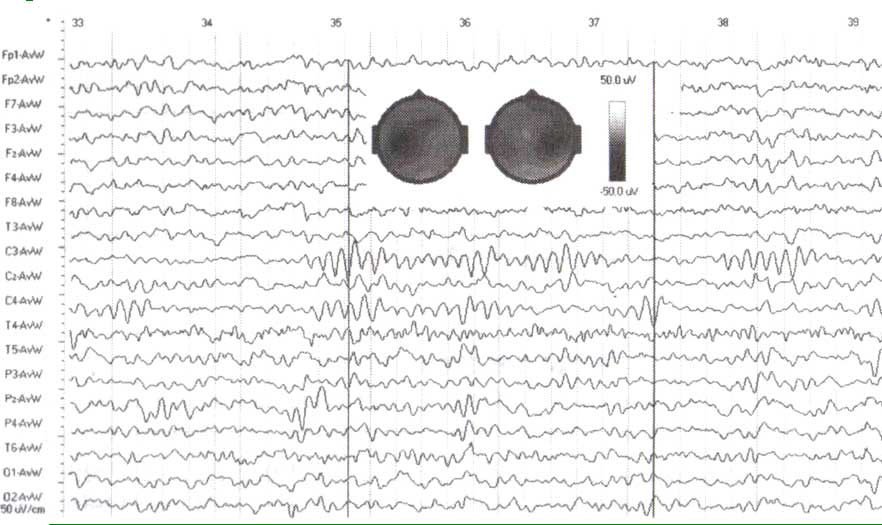
ЭЭГ, зарегистрированная у здорового взрослого субъекта в течение спокойного бодрствования с открытыми глазами. Обратите внимание на различные ритмы с частотой приблизительно 10 циклов в секунду в отведениях С3 и С4. Вставка — две топограммы потенциалов, снятых в два момента времени, указанных черными вертикальными линиями.
Сенсомоторные ритмы имеют немного более высокую частоту, чем затылочные альфа-ритмы. На рис. 1 изображен фрагмент ЭЭГ здорового взрослого человека в состоянии с открытыми глазами. Можно ясно видеть два различных ритмичных паттерна ЭЭГ в отведениях СЗ и С4. Эти колебания, по-видимому, независимы друг от друга и имеют специфическую форму с острыми отрицательными пиками, которые напоминают греческую букву μ (мю). Эта определенная форма дает одно из названий этого ЭЭГ-паттерна — мю-ритма.
Активируется µ-ритм во время умственной нагрузки и психического напряжения. µ-ритм снижается (или в небольшом числе случаев нарастает) по амплитуде при двигательной активации или соматосенсорной стимуляции, в связи с чем его еще называют «сенсоримоторным ритмом», ритм локализован над сенсомоторными участками коры (см. также карты потенциалов, построенные для двух различных моментов времени). В некоторых статьях этот ритм также называют роландическим, потому что он обнаруживается над роландической бороздой. Сенсомоторные ритмы могут блокироваться движением соответствующей руки или прикосновением к руке. И наоборот, расслабление мускулов увеличивает сенсомоторные ритмы.
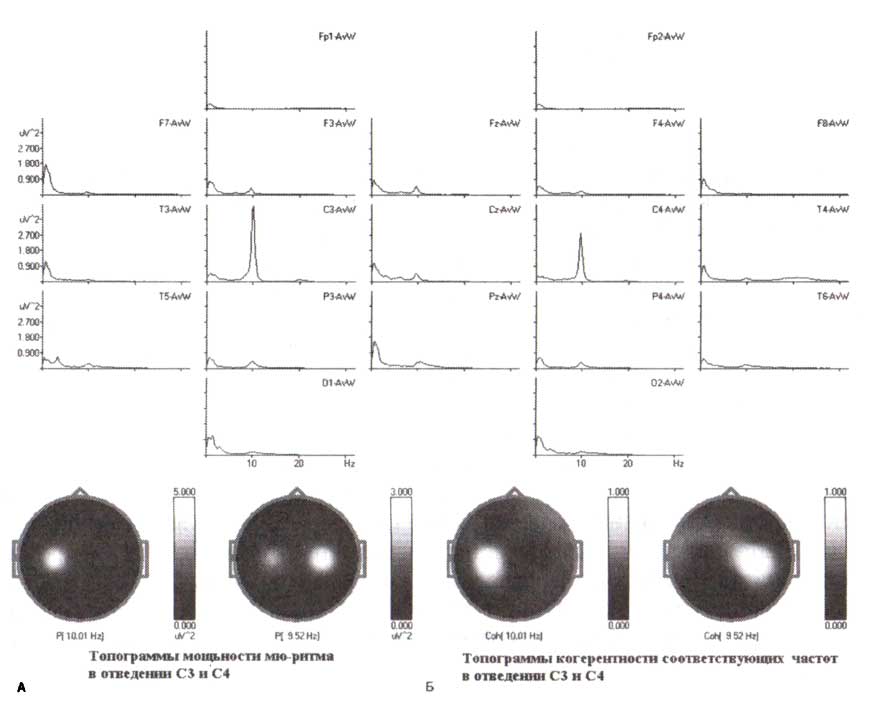
Пример спектров мощности здорового взрослого субъекта. А. Топограммы распределения мощности левого и правого мю-ритмов в максимуме их амплитуды. Обратите внимание, что эти мю-ритмы немного отличаются по частоте. Б. Топограммы когерентности отведений СЗ и С4 соответственно. Обратите внимание, что положительный пик когерентности в одной стороне сопровождается отрицательным пиком в противоположной стороне, что указывает на независимость левых и правых мю-ритмов.
Здесь надо подчеркнуть, что хотя мю-ритмы обнаружены и в левом, и в правом полушариях (и во многих случаях выглядят весьма симметричными в спектрах ЭЭГ), они обычно независимы, то есть производятся независимыми генераторами. Это продемонстрировано на рис. 2 при помощи когерентного анализа.
На рис. 2 представлены спектры, вычисленные для здорового человека, во всех 19 каналах в состоянии спокойного бодрствования с открытыми глазами. Как выявляется на спектрах ЭЭГ, в ЭЭГ доминируют два различных ритма: один с максимумом мощности ЭЭГ в отведении СЗ с пиковой частотой 10 Гц и другой с максимумом мощности ЭЭГ в отведении С4 с пиковой частотой 9,5 Гц. Карты когерентности, вычисленной отдельно для СЗ и С4, представлены внизу справа. Как видно, сенсомоторный ритм с максимумом в СЗ имеет самую высокую когерентность на частоте левого ритма (10 Гц) с близлежащими электродами левого полушария и самую низкую когерентность с С4 электродом и, наоборот, ритм в С4 не работает синхронно с ритмом в отведении СЗ на своей пиковой частоте (9,5 Гц).
Поскольку он обычно имеет низкую амплитуду и при визуальном анализе в большинстве случаев неотличим от альфа-ритма, он не имеет большого клинического значения. Однако применение компьютерных методов анализа с использованием цифровой фильтрации, усреднения, поперечных биполярных отведений и функциональных двигательных проб позволяет регулярно регистрировать его у всех пациентов, а его специфическая привязанность к области сенсомоторной коры делает его особенно чувствительным к нарушениям кровообращения в русле средней мозговой артерии, питающей эти отделы мозга1.
Реактивность мю-ритма ЭЭГ при восприятии речи у детей
Еще в первой половине ХХ века Л.С. Выготский утверждал, что мышление ребенка развивается в зависимости от овладения социальными средствами мышления, т.е. в зависимости от речи2. В настоящее время признается, что развитие речевой сферы является важнейшим фактором, определяющим когнитивное и социальное развитие ребенка. Исходя из тесной связи между развитием речи и мышления, размер активного словаря ребенка, т.е. количество слов, которое он понимает и произносит, расценивают как непрямой показатель уровня его вербального интеллекта3.
При регистрации ЭЭГ и МЭГ у взрослых во время восприятия целостных сообщений, состоящих из одного или нескольких предложений, было установлено участие в понимании речи нейрофизиологических процессов, отражающихся в росте мощности тета-, бета- и гамма-ритмов456, росте7 или падении альфа-8, а также альфа- и мю-ритмов9. У детей младшего возраста (8—12 мес) и дошкольников (4—7 лет) при социальном взаимодействии с взрослым, включающем прослушивание сказок, песен и рассказов, при открытых глазах росла мощность ЭЭГ в низкочастотном диапазоне (3.6—5.6 Гц у младшей группы и 4.0—7.6 Гц — у старшей)10. При этом значения мощности разновидности альфа-активности — мю-ритма (6.4—8.4 Гц у младшей группы и 8.4— 10.4 Гц — у старшей) — снижались. Такие конкурентные изменения указанных ритмов ЭЭГ авторы исследования связывали с ростом когнитивной и эмоциональной нагрузки у детей в ситуации социального взаимодействия. Согласно единичным исследованиям детей в возрастном периоде от полутора до трех лет11, т.е. именно в том возрасте, когда активно осваивается восприятие предложений12, показано, что у малышей 18 и 24 мес в условиях записи ЭЭГ при открытых глазах восприятие предложений, обозначающих действия, сопровождается десинхронизацией диапазона 6—10 Гц в центральных и затылочных областях мозга. Несколько иные результаты получены в недавних исследованиях процессов слухового восприятия речи у взрослых и детей 10—12 лет1314. В ответ на предъявление аудиозаписей корректных предложений и записей, содержащих ошибки, у испытуемых обеих групп в состоянии открытых глаз выявлено увеличение амплитуды не только в стандартных тета- и бета-диапазонах ритмов ЭЭГ, но также — в центральных регионах — низкочастотного альфа-ритма (8.0—10.9 Гц). Однако, когда авторы представляют результаты исследования тета- и бета-ритмов, нельзя исключить, что изменения амплитуды этих частотных диапазонов ЭЭГ вполне могут отражать изменения тонического напряжения мышц скальпа, связанное с ментальной активностью15. Установлено, что ЭМГ мышц скальпа когерентны с мощностью в стандартных тета- и бета-диапазонах и реципрокны мощности высокочастотного альфа-диапазона16171819.
Одним из приемов для изучения нейрофизиологических особенностей восприятия речи на основе анализа паттерна ЭЭГ является методика, описанная C. Krause и соавт.20, которая заключается в следующем: предъявляются два вида стимулов, один из которых представляет нативную запись речи, а другой — такую же запись, но воспроизведенную в реверсном режиме. Этот второй стимул является речеподобным сигналом, лишенным семантического содержания. Таким образом, применение приема, включающего сравнение изменений ЭЭГ в ответ на нативную и реверсированную речь, позволяет оценить особенности активации ЦНС испытуемых в ответ на осмысленные сообщения, доступные для их понимания.
Читайте также
Footnotes
- Pfurtscheller G. m-Rhytmen: Ursprung, Reaktivitat, Ableitung und klinische Bedeutung. EEG-Labor.-1986, Bd. 8, H. 2, s. 46-57.
- Выготский Л.С. Мышление и речь. М., Л.: Государственное социально-экономическое изд-во, 1934. 324 с.
- Cristia A., Seidl A., Junge C., Soderstrom M., Hagoort P., Predicting individual variation in language from infant speech perception measures. Child Dev. 2014. 85: 1330-1345.
- Ding N., Melloni L., Zhang H., Tian X., Poeppel D. Cortical tracking of hierarchical linguistic structures in connected speech. Nature Neuroscience. 2016. 19 (1): 158-164.
- Lewis A.G., Schoffelen J.M., Schriefers H., Bastiaansen M. A predictive coding perspective on beta oscillations during sentence-level language comprehension. Front. Hum. Neurosci. 2016. 10: 85. eCollection 2016.
- Mai G., Minett J.W., Wang W.S. Delta, theta, beta, and gamma brain oscillations index levels of auditory sentence processing. NeuroImage. 2016. 133: 516-528.
- Meyer L., Obleser J., Friederici A.D. Left parietal alpha enhancement during working memory-intensive sentence processing. Cortex. 2013. 49 (3): 711-721.
- Lam N.H., Schoffelen J., Udden J., Hulten A., Hagoort P. Neural activity during sentence processing as reflected in theta, alpha, beta, and gamma oscillations. NeuroImage. 2016. 142: 43-54.
- Saltuklaroglu T., Bowers A., Harkrider A.W., Casenhis-er D., Reilly K.J., Jenson D.E., Thornton D. EEG mu rhythms: rich sources of sensorimotor information in speech processing. Brain Lang. 2018. 187: 41-61.
- Orekhova E.V., Stroganova T.A., Posikera I.N., Elam M. EEG theta rhythm in infants and preschool children. Clin. Neurophysiol. 2006. 117 (5): 10471062.
- Antognini K., Daum M. Toddlers show sensorimotor activity during auditory verb processing. Neuro-psychologia. 2017.
- Kuhl P.K. Brain mechanisms in early language acquisition. Neuron. 2010. 67 (5): 713-727.
- Schneider J.M., Abel A.D., Ogiela D.A., Middleton A.E., Maguire M.J. Developmental differences in beta and theta power during sentence processing. Dev. Cogn. Neurosci. 2016. 19: 19-30.
- Schneider J.M., Abel A.D., Ogiela D.A., McCord C., Maguire M.J. Developmental differences in the neural oscillations underlying auditory sentence processing in children and adults. Brain Lang. 2018. 186: 17-25.
- Cacioppo J.T., Martzke J.S., Petty R.E., Tassinary L.G. Specific forms of facial EMG response index emotions during an interview: from Darwin to the continuous flow hypothesis of affect-laden information processing. J. Pers. Soc. Psychol. 1988. 54 (4): 592-604.
- Goncharova I.I., McFarland D.J., Vaughan T.M., Wol-paw J.R. EMG contamination of EEG: spectral and topographical characteristics. Clin Neurophysiol. 2003. 114 (9):1580-1593.
- Wijsman J., Grundlehner B., Liu H., Hermens H., Penders J. Towards mental stress detection using wearable physiological sensors. Conf. Proc. IEEE Eng. Med. Biol. Soc. 2011: 1798-1801.
- Fitzgibbon S.P., DeLosAngeles D., Lewis T.W., Powers D.M., Grummett T.S., Whitham E.M., Ward L.M., Willoughby J.O., Pope K.J. Automatic determination of EMG-contaminated components and validation of independent component analysis using EEG during pharmacologic paralysis. Clin Neurophysiol. 2016. 127 (3): 1781-1793.
- Janani A.S., Grummett T.S., Lewis T.W., Fitzgibbon S.P., Whitham E.M., DelosAngeles D., Bakhshayesh H., Willoughby J.O., Pope K.J. Evaluation of a minimum-norm based beamforming technique, sLORETA, for reducing tonic muscle contamination of EEG at sensor level. J. Neurosci. Methods. 2017. 288: 17-28.
- Krause C., Porn B., Lang H. Relative alpha desynchronization and synchronization during speech perception. Brain Res. 1997. 5 (3): 295-299.