С позиций нормальной физиологии нервная система рассматривается как возбудимая ткань: она способна менять трансмембранную разницу потенциалов при раздражении, а в некоторых случаях — спонтанно. Отдельно взятый нейрон находится в непрерывном процессе биоэлектрогенеза.
Регистрация активности нейронов показывает непрерывную смену базовых электрофизиологических процессов: потенциал покоя, потенциал действия, постсинаптические потенциалы (ПСП): возбуждающий или тормозный. Других источников электрического поля, кроме нейронов, в головном мозге нет.
Классификация нейронов
По структуре
Полярность

Большинство нейронов можно подразделить на:
- Униполярные (англ. unipolar neuron) – нейроны с одним отростком
- Биполярные (англ. bipolar neuron) – имеют один аксон и один дендрит
- Мультиполярные (англ. multipolar neuron) -имеют один аксон и несколько дендритов
- Клетка Гольджи I – нейроны с длинными аксональными отростками
- Клетка Гольджи II – нейроны либо не имеющие аксона, либо имеющие короткий аксон
- Псевдоуниполярные (англ. pseudounipolar neuron) – имеет один отросток, который является и дендритом и аксоном
- Нейроны без аксона (англ. anaxonic neuron) – нейрон, у которого нет аксона, либо его сложно отлечить от дендритов
Другие
Некоторые нейроны могут быть идентифицированы согласно их местоположению и структурным особенностям. Вот некоторые примеры:
- Корзинчатый нейрон (англ. basket cell) – интернейроны, которые образуют плотное сплетение терминалей вокруг сомы клеток-мишеней, обнаруживаются в коре и мозжечке;
- Клетки Бетца (англ. Betz cell) – крупные двигательные нейроны;
- Клетки Лугаро (англ. Lugaro cell) – интернейроны мозжечка;
- Средние шипиковые нейроны (англ. medium spiny neuron) – составляют полосатое тело;
- Клетки Пуркинье (англ. purkinje cell) – крупные нейроны, расположенные мозжечке, относятся к мультиполярным клеткам Гольджи I;
- Пирамидные клетки (англ. pyramidal cell) – нейроны с треугольной сомой, тип Гольджи I;
- Клетки Реншоу (англ. renshaw cell) – нейроны, связанные обоими концами с альфа-моторными нейронами;
- Униполярные микроворсинчатые (щеточные) клетки (англ. unipolar brush cell) – интернейроны с уникальным дендритом, заканчивающимся кистевидным пучком;
- Гранулярные клетки (англ. granule cell) – тип нейрона Гольджи II;
- Веретенообразные нейроны (англ. spindle neurons) – интернейроны, которые соединяют разные области мозга.
По функциональной активности
- Афферентные нейроны передают информацию от тканей и органов в центральную нервную систему и также называются сенсорными нейронами .
- Эфферентные нейроны (двигательные нейроны) передают сигналы от центральной нервной системы эффекторным клеткам.
- Интернейроны соединяют нейроны в определенных областях центральной нервной системы.
Афферентный и эфферентный также обычно относятся к нейронам, которые, соответственно, передают информацию или отправляют информацию от мозга.
Пирамидные нейроны как простейшие электрические диполи
Маленькие электрические токи всех дендритов одного нейрона суммируются по-разному в различных типах нейронов. Если нейрон — звездчатая клетка и возбуждающие синапсы покрывают множество дендритов, ориентированных в различных направлениях, суммарный ток на уровне клетки в целом будет близок к нулю, и суммарный потенциал, произведенный звездчатой клеткой, будет незначителен. Но если нейрон — пирамидная клетка и возбуждающие синапсы расположены на апикальных дендритах, суммарный ток пирамидной клетки будет направлен вдоль апикального дендрита, и нейрон может быть представлен как маленький диполь с отрицательным полюсом в апикальном дендрите и с положительным полюсом в базальной части нейрона (рис. 1). Также, если возбуждающие синапсы активизированы на соме и базальных дендритах нейрона, нейрон становится диполем, который заряжен отрицательно в соме и положительно — в апикальном дендрите.
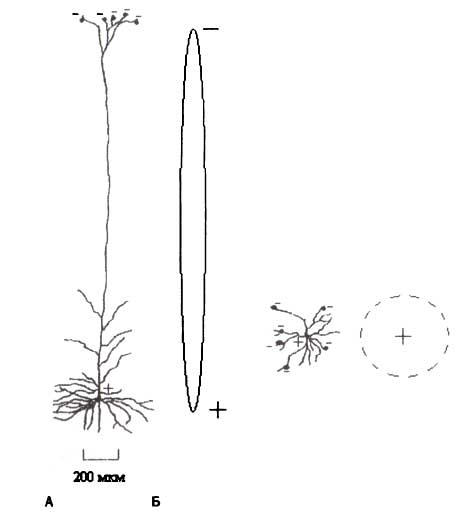
А. Пирамидная клетка с длинным апикальным дендритом. Б. Звездчатая клетка. На этом примере поток Na* внутрь клетки вызван связыванием молекул возбуждающего медиатора глутамата с рецепторами дендрита. Обратите внимание, что ионные токи в пирамидной клетке формируют отдельный диполь, в то время как ионные токи в звездчатой клетке формируют вне клетки нулевое поле.
ЭЭГ — это результат общей электрической активности пирамидных нейронов, то есть суммирования сотен тысяч маленьких диполей, соответствующих этим пирамидным нейронам. Диполи образованы локальными токами, которые связаны с возбуждающими и тормозными постсинаптическими потенциалами. Возбуждающие постсинаптические потенциалы деполяризуют мембрану, приближая ее к порогу возникновения спайка, и делают выше вероятность овершута (перезарядки мембраны при возникновении ПД). Тормозные постсинаптические потенциалы гиперполяризуют мембрану, уводя ее от порога генерации спайка, и таким образом уменьшают вероятность перезарядки.
Продолжительность постсинаптических потенциалов в десять раз больше продолжительности отдельного спайка. Как мы знаем, продолжительность постсинаптического потенциала зависит от типа соединения рецептора-медиатора, но длится по крайней мере 20—30 мс, что соответствует частоте 30—50 Гц. Продолжительность спайка — приблизительно 1 мс, что соответствует частоте 1000 Гц.
Кроме того, пространство, занятое синапсами, где производятся постсинаптические потенциалы, намного больше, чем пространство, занятое аксональным бугорком, где возникают спайки. Из этих двух факторов следует, что постсинаптические потенциалы играют критическую роль в генерации электрических потенциалов, регистрируемых со скальпа в диапазоне частот 0-70 Гц и известных как электроэнцефалограмма.
Дипольная структура внутри корковых потенциалов экспериментально демонстрируется, когда локальные электрические потенциалы, измеренные в различных корковых слоях посредством сетки микроэлектродов, помещенных в кору, показывают явную смену полярности потенциала в средних слоях коры. Эти внутрикорковые диполи производят скальповые потенциалы, которые могут быть вычислены согласно законам физики. Проблема вычисления скальповых потенциалов от известных внутрикорковых диполей известна как прямая задача.
Синхронизация нейронных элементов — другой фактор, который определяет скальповый электрический потенциал. Интуитивно понятно, что для того чтобы возник существенный потенциал на относительно отдаленном расстоянии от корковых слоев (на поверхности скальпа), нейроны должны быть не только должным образом ориентированы, но они должны работать вместе, синхронно. Например, область источника эпилептического спайка, который внезапно перекрывает фоновую электрическую активность головного мозга, была экспериментально оценена приблизительно в 2000 мм2, в то время как для других элементов ЭЭГ может быть достаточно кластера в 40—200 мм2. Так или иначе синхронизация нейронных элементов на расстоянии десятков миллиметров — необходимый фактор для возможности регистрации элементов ЭЭГ со скальпа.
Электрическая активность нейрона
Потенциал покоя
Мембрана нейрона за счет избирательной проницаемости для различных ионов и работы специальных устройств — «ионных насосов» — постоянно поддерживает разницу концентрации ионов1. В основном разница концентрации поддерживается для Na+ и К+: концентрация Na+ внутри клетки в 150 раз меньше, чем на ее поверхности. В результате между наружной и внутренней поверхностями мембраны образуется разность потенциалов — потенциал покоя (ПП). Введя микроэлектрод внутрь клетки, потенциал покоя можно измерить. В нервной клетке он составляет около -60 … —70 мВ, внутренняя поверхность мембраны заряжена отрицательно относительно наружной. Наличие разности потенциалов между внутриклеточной и внеклеточной средой обозначают термином «поляризация».
Потенциал покоя является тем базовым уровнем, от которого измеряются изменения поляризации мембраны в результате процессов возбуждения и торможения. Увеличение амплитуды потенциала покоя называется гиперполяризацией, а уменьшение — деполяризацией. Флюктуация уровня поляризации идет на мембране постоянно в основном в результате синаптического взаимодействия с другими нейронами.
Потенциал действия
Импульсная активность нейронов связана с процессом генерации потенциала действия (ПД). Потенциал действия возникает при снижении заряда мембраны до некоторого критического уровня — критического уровня деполяризации. В этот момент наступает электрический «пробой» мембраны: открываются ионные каналы, ток Na+ устремляется внутрь клетки. В результате лавинообразного тока ионов генерируется ПД. Потенциал действия имеет очень высокую амплитуду по отношению к исходному уровню поляризации. Уровень ПП составляет — 60 мВ, а при генерации ПД заряд мембраны меняется на противоположный (овершут) и достигает порядка +70 … +120 мВ. ПД имеет длительность порядка 1—2 мс. Восстановление исходного уровня поляризации участка мембраны, на котором произошла генерация ПД, обеспечивается работой ионных насосов, которые восстанавливают исходный уровень градиентов концентрации ионов. В этот период мембрана становится невозбудимой — рефрактерной и генерация следующего потенциала действия невозможна.
Потенциал действия подчиняется закону «все или ничего»: он либо генерируется при достижении критического уровня деполяризации мембраны, либо не возникает, если уровень локального изменения поляризации недостаточен. Во втором случае наблюдаются градуальные электрические реакции, которые представляют собой ПСП. В зависимости от вида медиатора выделяют возбуждающие и тормозные синапсы. Типичными возбуждающими синапсами являются, например, холинергические, адренергические. Основным тормозным медиатором в ЦНС выступает гамма-аминомасляная кислота (ГАМК). При выделении в синаптическую щель возбуждающего медиатора на постсинаптической мембране возбуждающего синапса происходит градуальная деполяризация — генерируется возбуждающий постсинаптический потенциал (ВПСП). В тормозном синапсе происходит гиперполяризация мембраны, соответственно, генерируется тормозный постсинаптический потенциал (ТПСП). ПСП отличаются от потенциала действия локальностью возникновения, декрементным (затухающим) распространением по мембране, сравнительно малой амплитудой (не более 20—40 мВ), а главное — большой длительностью (до 20—50 мс). Перечисленные свойства ПСП обеспечивают возможность суммации градуальных потенциалов во времени и пространстве, обеспечивая процессы интеграции на уровне нейрона.

Итак, на уровне нейрона можно выделить два вида электрических процессов: генерация импульсной активности (потенциал действия) и градуальные изменения поляризации (ПСП). Как следует из изложенного выше, ПД представляет собой значительно более редкое событие, чем градуальные потенциалы. Приблизительное соотношение между вероятностью этих событий можно получить из следующего сопоставления. Средняя частота генерации ПД нейроном составляет около 10 в секунду. Но по каждому из синаптических окончаний к дендритам и соме нейрона притекает соответственно, в среднем, 10 синаптических воздействий за секунду. На поверхности дендритов и сомы одного коркового нейрона могут оканчиваться до нескольких тысяч синапсов. В результате объем синаптической бомбардировки одного нейрона, а соответственно, и вероятность возникновения градуальных реакций, превосходит вероятность генерации ПД в несколько десятков тысяч раз. Кроме того, длительность ПСП в 50—100 раз превышает длительность ПД.
Таким образом, перечисленные основания позволяют считать, что основу суммарной ЭЭГ составляют соматодендритные ПСП нейронов коры.
Ритмическая активность нейронов
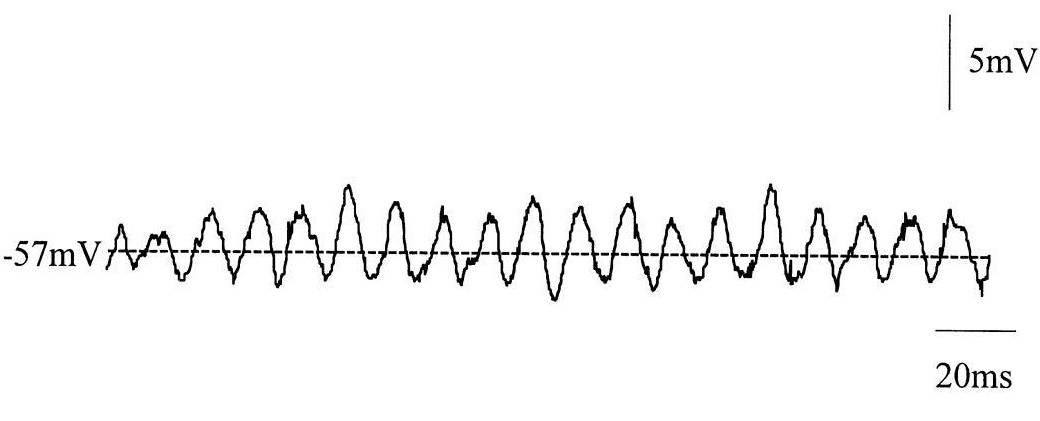
Ритмическая активность на микроуровне вызвана генерацией потенциала действия, в результате изменения заряда мембраны. Нейроны генерируют множественные последовательные потенциалы – пачки спайков (spike trains), которые являются основой для нейронного кодирования и передачи информации в мозге. Они формируют все виды паттернов.2 Помимо стандартной колебательной активности одиночные нейроны способны генерировать регулярный паттерн подпороговых колебаний мембранного потенциала (англ. regular pattern of SMPO), что не сопровождается формированием потенциала действия. Это происходит, когда постсинаптический потенциал не смог достигнуть порога возбудимости нейрона и не активировал его, либо это может быть связано со спонтанными изменениям потенциала нейронов. Характеризуется средней частотой 86 Гц и средней амплитудой 3,3 мВ.
На мезоуровне происходит взаимодействие больших нейронных популяций между собой через связи головного мозга, здесь же в коре головного мозга обрабатывается сенсорная информация, которая вовлекает в себя подкорковые структуры, для чего необходима тесная связь между этими областями. Популяция нейронов может запускаться вместе либо реагируя на входной перцептивный или выходной моторный импульс, либо из-за синаптических взаимодействий. Благодаря синаптическим взаимодействиям паттерны возбуждения различных нейронов синхронизируются, что вызывает изменение ритма биоэлектрической активности мозга, при этом если импульс пришел в одну фазу, то сигнал усиливается, а если в разные, то наоборот ослабевает. Синхронизированные паттерны импульсов одной нейронной популяции приводит к синхронизации других областей коры, что вызывает высокоамплитудные колебания потенциала локального поля (LFP). Синхронная деятельность нейронов является главным условием для формирования колебательной активности мозга. В ином случае, если одна популяция нейронов отправит другой недостаточной силы импульс, либо он придет, когда часть нейронов еще находится в рефрактерном периоде, то вторая популяция не сможет поддержать возбуждение, и волна в итоге затухает. Контролируется это эндогенно через локальные взаимодействия между возбуждающими и тормозными нейронами. В частности, тормозные интернейроны играют важную роль в создании нейронно-ансамблевой синхронии, создавая узкое окно для эффективного возбуждения и ритмически модулируя частоту испускания импульсов возбуждающих нейронов.3
Неритмическая активность нейронов
➥ Основная статья: Шум в биоэлектрической активности мозга
На уровне нейронов неритмическая активность может быть связана с молекулярными и синаптическими взаимодействими. Поток ионов через открытые ионные затворы вызывает 1/f-шум (также розовый шум, фликкер-шум или избыточный шум), что видно из спектральной плотности мощности (PSD) мембранного потенциала.
Регистрация активности нейронов
➥ Основная статья: Регистрация активности нейронов
Одним из самых распространенных методов регистрации активности нейронов является электроэнцефалография. Связь ЭЭГ с элементарными электрическими процессами на уровне нейрона, безусловно, нелинейная. Предполагается, что ЭЭГ является результатом сложной суммации электрических потенциалов целостного нейронного ансамбля. Расчетным путем установлено, что активность, регистрируемая под одним электродом на поверхности головы, отражает суммацию постсинаптических потенциалов нейронов коры на площади 100—200 мм2 45.
Footnotes
- Электроэнцефалография : руководство / М. В. Александров, Л. Б. Иванов, С. А. Лытаев [и др.] / под ред. М. В. Александрова. — 3-е изд., перераб. и доп. — Санкт-Петербург: СпецЛит, 2020. — 224 с.
- Wang XJ (2010). “Neurophysiological and computational principles of cortical rhythms in cognition”. Physiol Rev. 90 (3): 1195–1268. doi:10.1152/physrev.00035.2008.
- Nunez PL, Srinivasan R (1981). Electric fields of the brain: The neurophysics of EEG. Oxford University Press.
- Бреже М. Электрическая активность нервной системы : пер. с англ. — М. : Мир, 1979. — 264 с.
- Кропотов Ю. Д. Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия : пер. с англ. — Донецк : Издатель Заславский А. Ю., 2010. — 512 с.