Значительное количество шума присутствует из-за колебаний и столкновений молекул. Этот вид шума называется тепловым (англ. thermal noise), он оказывает значительное влияние на функции ионных каналов нейронов. Ионный канал представляет собой макромолекулу, которая встроена в постоянно колеблющуюся среду, либо мембрану с фосфолипидами, либо внутриклеточные и внеклеточные растворы. Тепловые флуктуации, присутствующие в этих средах, вызывают многочисленные столкновения с ионным каналом, что приводит к спонтанным изменения конформации белка, некоторые из которых могут открыть или закрыть канал. Следовательно, ионный канал может открываться и закрываться стохастическим образом, такое явление называется канальным шумом (англ. channel noise).
Перенос ионов через биполярную мембрану при помощи ионофоров может вызвать берстовый шум (англ. burst noise) вызванный внезапными скачкообразными неустойчивыми переходами в системах с двумя или более дискретными уровнями напряжения или тока.
В центральных нейронах, и в частности в коре головного мозга, источником шума с наибольшей амплитудой является синаптический шум (англ. synaptic noise), который, как было установлено, является доминирующим во внутриклеточных записях in vivo. Синаптический шум описывает непрерывную и шумную «бомбардировку» центральных нейронов нерегулярными синаптическими импульсами. В частности, кора головного мозга in vivo характеризуется устойчивой и нерегулярной нейрональной активностью, которая в сочетании с очень высокой корковой взаимосвязью, ответственна за синаптическую активность в любом данном корковом нейроне, которая имеет решающее значение для его внутренних динамических свойств и реакций.
1/f шум
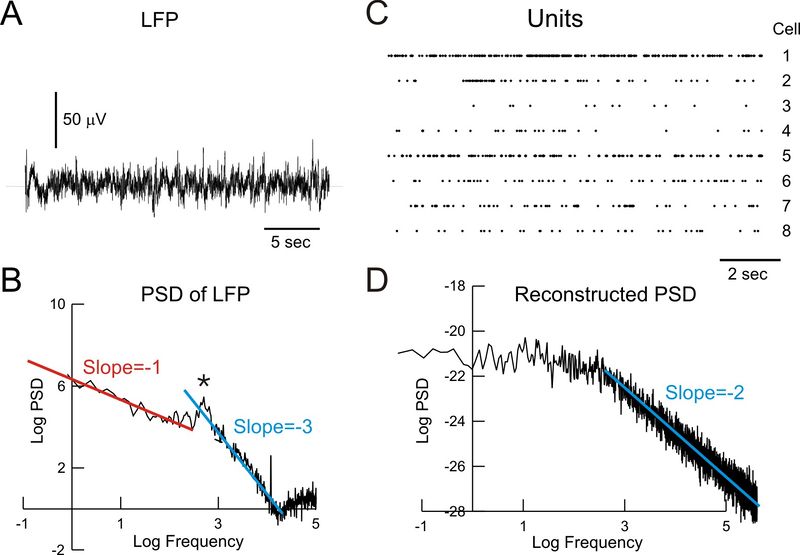
A. Запись LFP в теменной коре бодрствующей кошки B. Спектральная плотность мощности (PSD) LFP, показывающая две области масштабирования: 1/f на низких частотах (красная линия, наклон = -1) и 1/f³ на высоких частотах (синяя линия; наклон = -3). C. Активность одиночного нейрона, записанная с помощью системы из 8 вольфрамовых электродов. D. Попытка воссоздать сигнал LFP от активности одиночных нейронов. Низкочастотный конец PSD был постоянным (нулевой наклон), в то время как высокочастотный конец масштабировался как 1/f² (синяя линия, наклон = -2). Показатель -1 отсутствует.
1/f-подобный спектр мощности, регистрируемый на всех уровнях организации, указывает на аритмическую активность мозга, которая не содержит преобладающего временного масштаба (“безмасштабный”, “scale-free”). Спектр мощности потенциалов электрического поля мозга содержит преобладающий компонент «1/f»; то есть мощность имеет тенденцию падать с увеличением частоты в зависимости от степенной функции: P∞1/f ᵝ, где P – мощность, f – частота и β является параметром, обычно находящимся в диапазоне 0 ~ 3. Эта характеристика свободной от масштаба мозговой деятельности отличает ее от мозговых колебаний.1
На микроуровне существует ряд источников шума в нейронных мембранах, которые прямо или косвенно связаны с тепловым и канальным шумом, но изучаются отдельно. Поток ионов через открытые ионные затворы вызывает 1/f-шум (также розовый шум, фликкер-шум или избыточный шум), что видно из спектральной плотности мощности (PSD) мембранного потенциала. Эта миграция ионов через открытые и протекающие каналы и поры является источником другого типа шума, называемого дробовым шумом. Его происхождение можно найти в квантово-механических свойствах ионов и, в более общем смысле, участвующих частиц, в частности дискретности их электрического заряда. В отличие от упомянутого выше теплового шума, появление 1/f шума неизбежно связано с неравновесными состояниями рассматриваемой системы и, следовательно, играет роль в переходных и динамических состояниях.
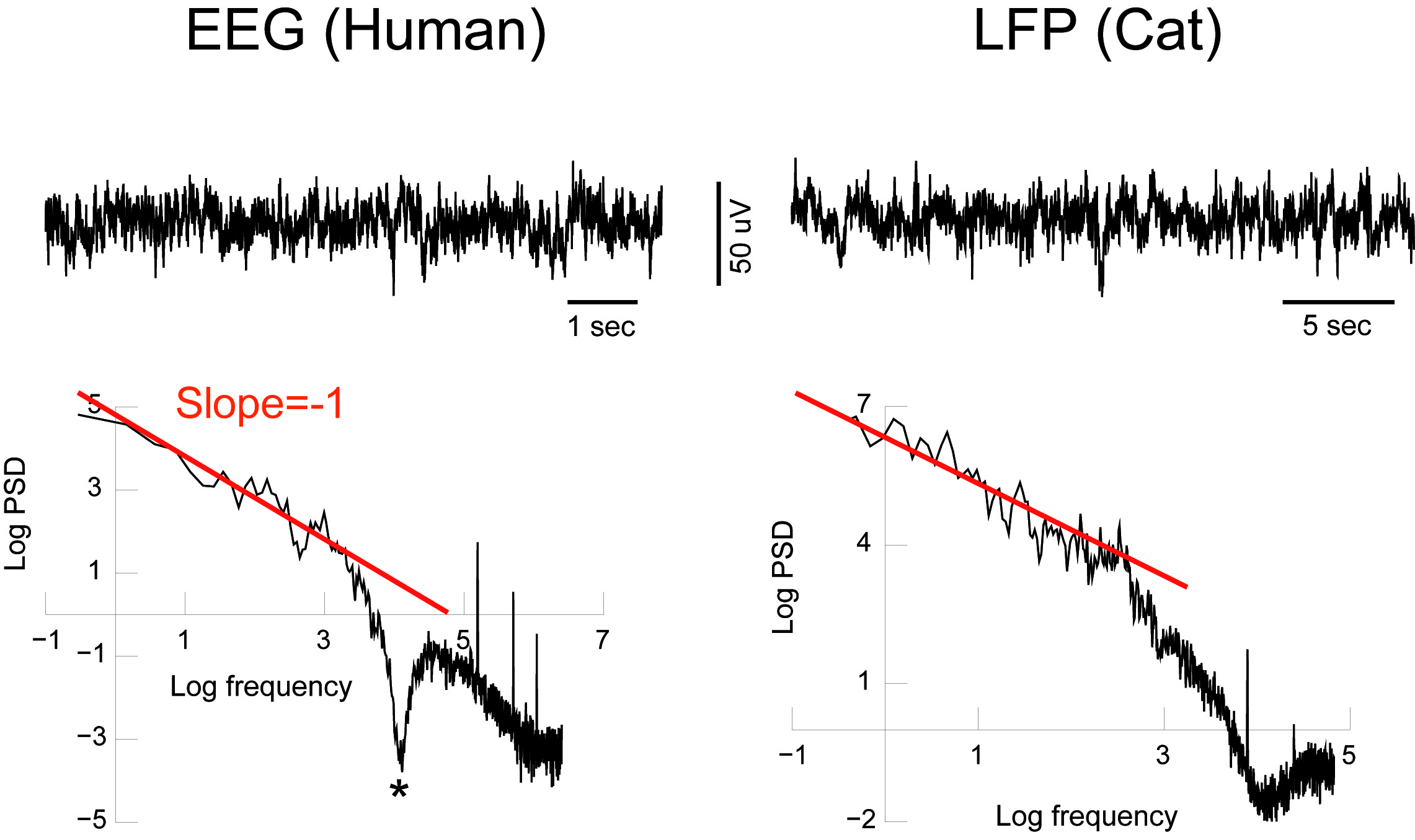
Кривые сверху показывают примеры записей ЭЭГ человека (слева) и записи LFP из теменной коры кошки (справа) во время бодрствования и сосредоточенном состоянии. Соответствующие спектры мощности (нижние) отображают приблизительное масштабирование 1/f на низких частотах. Прямые линии (красные) обозначают наклон -1. Сигналы не были отфильтрованы, за исключением режекторного фильтра на частоте 60 Гц для ЭЭГ и фильтров сбора данных на высоких частотах (не видимых в этом масштабе).
На мезоуровне одной из характеристикой сигналов LFP и ЭЭГ также является то, что они демонстрируют 1/f-подобный спектр мощности (“безмасштабный”, “scale-free”) на низких частотах, что показано в ряде исследований.23 Во время бодрствования и сна 1/f-шум может быть дополнительно проанализирован путем вычисления спектральной плотности мощности (PSD) LFP. Соответствующая PSD обычно отображает широкополосную структуру: во время бодрствования PSD показывает две различные области масштабирования в соответствии с полосой частот. Для низких частот (от 1 Гц до 20 Гц) PSD масштабируется приблизительно как 1/f, тогда как для более высоких частот (от 20 Гц до 65 Гц) масштабирование составляет приблизительно 1/f³. Во время медленного сна (англ. slow-wave sleep, SWS) дополнительная мощность на медленных частотах маскирует масштабирование 1/f, но такое же масштабирование 1/f³ присутствует и в высокочастотном диапазоне. Таким образом, эти результаты подтверждают, что масштабирование 1/f частоты, также присутствует в LFPs, но только во время бодрствования и для определенных диапазонов частот. Они также показывают, что глобальные переменные, такие как LFP, отображают широкополосную спектральную структуру, аналогичную стохастическим системам.
Спектры 1/f могут быть результатом самоорганизованных критических явлений, что позволяет предположить, что нейроны могут генерировать такую биоэлектрическую активность, но это утверждение противоречиво. Наконец, 1/f-шум может быть связан со способностью токов внеклеточной среды фильтровать часть спектра. К такому выводу пришли, отметив, что глобальная активность, реконструированная из многосайтовых записей, масштабируется идентично LFP, если предполагается «фильтр 1/f», и нет необходимости принимать самоорганизующиеся критические состояния в нейронной активности, Однако в последних исследованиях подчеркивалось, что 1/f-фильтр может быть необходим для объяснения результатов эксперимента.
На макроуровне 1/f-подобный спектр мощности (розовый шум),4 можно зарегистрировать при помощи неинвазивной электроэнцефалографии (ЭЭГ), магнитоэнцефалографии (МЭГ) и функциональной магнитно-резонансной томографии (фМРТ). Данная нейронная активность проявляется в норме и, предположительно, тесно связана как с когнитивной обработкой, так и с поддержанием функций мозга.5
Footnotes
- He BJ, et al. The temporal structures and functional significance of scale-free brain activity. Neuron. 2010;66:353–369. [PMC free article] [PubMed] [Google Scholar]
- Maimon G, Assad JA. Beyond Poisson: increased spike-time regularity across primate parietal cortex. Neuron. 2009;62:426–440. [PMC free article] [PubMed] [Google Scholar]
- Neuronal shot noise and Brownian 1/f2 behavior in the local field potential. Milstein J, Mormann F, Fried I, Koch C PLoS One. 2009; 4(2):e4338. [PubMed]
- Self-organized criticality: An explanation of the 1/f noise. Bak P, Tang C, Wiesenfeld K Phys Rev Lett. 1987 Jul 27; 59(4):381-384. [PubMed]
- Scale-free brain activity: past, present, and future. Trends In Cognitive Sciences. Biyu J He. May 3, 2014 [META]