Тета-ритм — ЭЭГ-активность от 4 до 8 Гц — был описан и систематизирован в работах 1950-х гг., т. е. последним из общепринятой ЭЭГ-ритмики.
Гиппокампальный тета-ритм — ритмическая активность в частотном диапазоне от 3 до 10 Гц, обнаруживаемая в гиппокампе млекопитающих и взаимосвязанных с ним анатомических структурах мозга.
Фронтальный тета-ритм средней линии — спонтанные или индуцированные выполнением задания кратковременные вспышки ритмической (от 5,5 до 8,5 Гц) активности, регистрируемые в области фронтальных отведений с максимумом в Fz. Наблюдается у здоровых взрослых испытуемых. Этот тета-ритм синхронизирован с реализацией действий в ответ на значимые события окружающей среды и ассоциируется с процессами вспоминания и кодирования следов памяти.
В классической нейрофизиологии вопрос об источнике и механизмах генерации активности тета-диапазона занимает особое место. Существует две модели, описывающие генерацию тета-активности. Первая (исторически) модель рассматривает гиппокамп как генератор тета-ритма. Вторая модель — таламическая, в соответствии с которой тета-ритм имеет единые с альфа-ритмом механизмы генерации. Согласно таламической модели, весь частотный диапазон от 4 до 13 Гц рассматривается как единый альфа-тета-континуум.
Нейронные механизмы
Связь с корковой активацией
В отличие от альфа-ритмов среднелобный тета-ритм может быть связан с активацией и увеличением метаболической активности в средней лобной области и передней поясной извилине. У пациентов с диагнозом эпилепсии электрическая стимуляция зон передней поясной извилины через интрацеребральные электроды приводила к возникновению 3—8 Гц ритма, регистрируемого в среднелобных зонах. Такая активность наблюдалась в среднелобных зонах у пациентов и без электрической стимуляции (Talairach и др., 1973). Исследовательская группа Гарвардского университета и университета Висконсина (США) (Pizzagalli и др., 2003) была первой, которая изучила связь между среднелобной тета-акгивностью и метаболизмом в человеческом мозгу. Одновременная регистрация электрической мозговой активности (ЭЭГ) и метаболизма глюкозы (ПЭТ-исследование) была выполнена на группе здоровых испытуемых. Данные ЭЭГ были проанализированы с позиций выявления мозгового источника генерации указанной активности, что позволило сопоставить ЭЭГ- и ПЭТ-данные на уровне оценки активации в вокселях. Именно для тета-ритма был выявлен самый большой кластер в передней поясной извилине (Поля Бродмана — 24/32), демонстрирующий положительную корреляцию между плотностью тока и метаболизмом глюкозы.
Связь с гиппокампальными тета-ритмами
Среднелобный тета-ритм в ЭЭГ человека часто связывается с гиппокампальной тета-активносгью в мозге млекопитающих. Причина такого связывания очень проста: гиппокамп млекопитающих обладает поразительной способностью к генерации тета-ритма. Эта ритмическая активность может быть зарегистрирована макроэлектродами во всех частях гиппокампа и в большинстве связанных с ним анатомических структур. Этот тип общего внеклеточно регистрируемого явления отражает кооперативное поведение большого количества гиппокампальных пирамидных клеток. Гиппокампальные пирамидные клетки, имеющие одинаковую ориентацию в пространстве, генерируют синхронную и периодическую электрическую активность в частоте тета-ритма. Частоты этих колебаний изменяются в весьма широком диапазоне от 4 до 10 Гц, имеют свои особенности и зависят от функционального состояния. Эти типы колебаний обычно объединяются под общим названием «гиппокампальные тета-ритмы» или, в более широком смысле, «лимбические тета-ритмы».
Лимбическая система гиппокампальных тета-ритмов
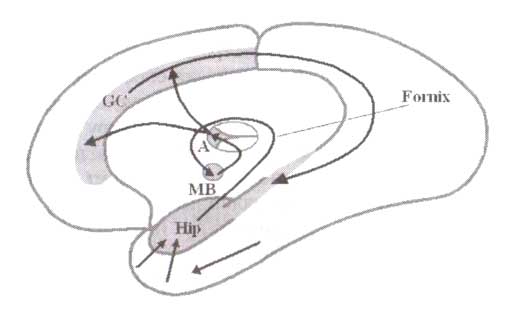
Гиппокамп (Hip) получает мультимодальные сенсорные входы, смешанные со входами от передней поясной коры, обрабатывает их и посылает результаты сосцевидным (мамиллярным) телам (МВ) гипоталамуса через путь, названный Сводом (Fornix), где они объединяются с другими входами и посылаются к переднему ядру (А) таламуса. Переднее ядро служит релейным ядром, посылающим свои выходы к поясной извилине (GC).
Мозговые структуры, в которых возможна регистрация тета-ритма, изображены на рис. 1. Эти структуры фактически представляют собой петлю (круг). Информационный поток в этом круге может быть сведен к следующему: 1) гиппокамп получает полисенсорный сигнал через обонятельную кору средней височной доли от различных височных и теменных сенсорных областей и сигнал от передней части поясной извилины; 2) гиппокамп интегрирует эту информацию и посылает результаты обработки к мамиллярным телам через гипоталамус, через пучок аксонов, называемый сводом (fornix); 3) мамиллярные тела посылают информацию дальше к переднему ядру таламуса через мамиллярноталамический пучок; 4) переднее ядро таламуса перенаправляет информацию к передней поясной извилине, а ее обратные связи с гиппокампом замыкают петлю Папеца.
Типы тета-ритма
В 1975 году Крамис и др. сделали предположение, что у крыс существует два типа гиппокампального тета-ритма, характеризующегося различными поведенческими и фармакологическими свойствами. По их мнению, тета первого типа («устойчивая к атропину») появляется во время движения и других вариантах «произвольного» поведения, а также во время быстрого сна, имеет частоту примерно около 8 Гц и не подвержена влиянию атропина. Тета второго типа («чувствительный к атропину») появляется во время неподвижности и во время анестезии, вызванной уретаном, имеет частоту в диапазоне 4-7 Гц и устраняется при введении атропина.1
Многие более поздние исследования подтвердили общую концепцию, согласно которой гиппокампальный тета-ритм может быть разделена на два типа, хотя возникали споры относительно свойств каждого из типов. Тета второго типа сравнительно редко встречается у крыс без анестезии: ее можно обнаружить на короткое время, когда животное готовится сделать движение, но еще не выполнило его, но сообщалось только о продолжительных периодах у животных, которые находятся в состоянии замороженной неподвижности из-за присутствия поблизости хищника, такого как кошка или хорек.2
Аномальные тета-ритмы
Клиническое значение нормального среднелобного тета-ритма и теменного альфа-тета-ритмов ограничено по нескольким причинам. Во-первых, эти ритмы имеют короткую длительность и появляются только в состояниях выполнения каких-либо заданий в зависимости от трудности задачи. Во-вторых, даже при выполнении заданий они могут быть замечены в спектрах ЭЭГ в форме пиков только у части здорового населения. В-третьих, трудность регистрации этих ритмов создает проблемы для систематических исследований их нейронных механизмов.
Подтип среднелобного тета-ритма при синдроме нарушения внимания и гиперактивности
На текущей стадии исследований мы имеем представления о том, что существует условие, связанное с отклонениями от нормы при генерации среднелобного тета-ритма. Это редкий подтип — часть населения с синдромом нарушения внимания и гиперактивности. Особыми признаками такого населения являются сложные социальные отношения и неспособность корректировать свое поведение. С электрофизиологической точки зрения этот СНВГ-подтип характеризуется необычной выраженностью среднелобного тета-ритма. Присутствие ритма выражено в чрезвычайно высоком и остром пике на спектрах ЭЭГ в Fz-отведении в диапазоне частот от 5,5 до 8 Гц и чрезвычайно низкой синхронизацией тета-ритма в ответ на значащие стимулы при выполнении двухстимульной GO/NOGO и математических задач.
Тета-ритмы в нелобных областях
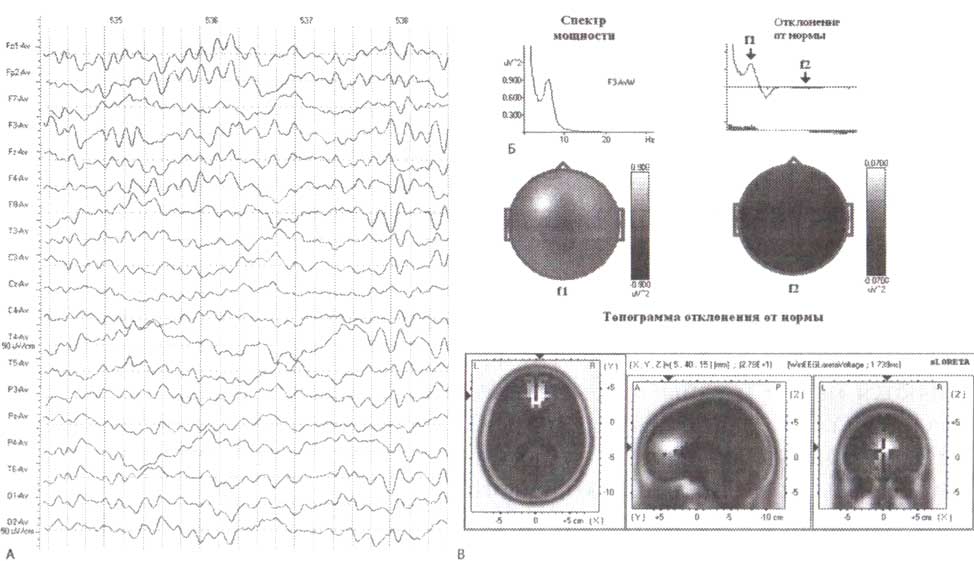
А. Фрагмент ЭЭГ, зарегистрированной у 32-летнего пациента через 5 дней после внутричерепной мозговой травмы. Б. Спектр мощности сигнала в отведении F3 и отклонение от нормы в отведении F3. С. Карты отклонений от нормы в тета- и бета-частотах и изображения sLORETA эквивалентных генераторов тета-ритма.
Появление тета-ритмов в других областях кроме отведения Fz (и до некоторой степени в Pz) следует считать ненормальным. В таких случаях необходимо сравнение с нормативной базой данных для того, чтобы определить уровень достоверности наблюдаемого отклонения от нормы (рис. 2). ЭЭГ была зарегистрирована на пациенте через нескольких дней после закрытой черепно-мозговой травмы. Можно видеть высокий по амплитуде тета-ритм в F3-, Р4-отведениях, мощность которых значительно (р < 0,001) отличается от нормативных данных. Обратите внимание также на то, что отклонения в диапазоне тета-ритма сопровождаются чрезвычайно низкой мощностью бета-активности, широко распределенной по коре.
Заключение
Тета-ритм — ритм с частотой больше 4 и меньше 8 Гц. Японские ученые впервые индуцировали проявление тета-активности ЭЭГ, предложив испытуемым умственную задачу на непрерывное арифметическое сложение. Эта тета-активность проявилась в виде последовательности ритмичных волн с частотой 6—7 Гц с максимальной амплитудой вокруг лобной средней линии и была названа среднелобным тета-ритмом. Среднелобный тета-ритм часто связывается с гиппокампальными тета-ритмами. Существуют гипотезы, согласно которым гиппокампальные тета-колебания вовлечены в кодирование следов памяти и извлечение из памяти. Регистрация электрической активности в гиппокампе человека доступна только в редких случаях сте-реотактических операций у пациентов с эпилепсией со вживленными для диагностических и лечебных целей электродами. Среднелобный тета-ритм указывает на индивидуальные различия и связан с некоторыми чертами индивидуальности: выраженность среднелобного тета-ритма отрицательно коррелирует с уровнем тревожности и невротизма личности, положительно — с уровнем экстраверсии. Среднелобный тета-ритм также коррелирует с изменениями уровня тревожности, вызванными успокаивающими лекарственными препаратами.
Читайте также
- Гиппокампальный тета-ритм
- Среднелобный тета-ритм
- Тета-ритм в процессах формирования памяти
- Тета-ритм при решении задач
- Генерация тета-ритма
- Тета (θ)-ритм и его происхождение
- тета-Активность
Footnotes
- Kramis R, Vanderwolf CH, Bland BH. Two types of hippocampal rhythmical slow activity in both the rabbit and the rat: relations to behavior and effects of atropine, diethyl ether, urethane, and pentobarbital. Exp Neurol. 1975 Oct;49(1 Pt 1):58-85. doi: 10.1016/0014-4886(75)90195-8.
- Sainsbury RS, Heynen A, Montoya CP. Behavioral correlates of hippocampal type 2 theta in the rat. Physiol Behav. 1987;39(4):513-9. doi: 10.1016/0031-9384(87)90382-9.