Быстрые трансмиттеры
Основу процессов обработки информации мозгом составляет преобразование импульсной активности пресинаптического нейрона в более медленный мембранный потенциал постсинаптического нейрона. В свою очередь, информационные процессы могут быть подразделены на два класса операций нейронных сетей: информационный поток и информационная модуляция. Реализация этих операций обеспечивается с помощью нейромедиаторов двух разных классов: медиаторов быстрого и медленного действия. Они выполняют разную функциональную роль в обеспечении процессов обработки информации.
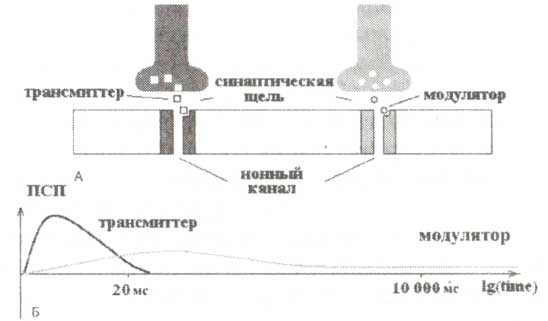
А. Схематически представлено действие двух типов нейромедиаторов. В обоих случаях высвобождение медиатора в синаптическую щель вызывается спайном, приходящим на пресинаптическую терминаль. Как нейротрансмиттер, так и нейромодулятор действуют на соответствующие каналы мембраны, что приводит к их открытию. При этом действие нейромедиатора по времени более длительно по сравнению с нейротрансмитерром. Б. График временной динамики постсинаптических потенциалов, индуцируемых нейротрансмиттером и нейромедиатором. Обозначения: ПСП — постсинаптический потенциал.
Информационные потоки поддерживаются быстрыми медиаторами (рис.1). Благодаря тому что эти медиаторы могут за несколько миллисекунд связываться с соответствующими рецепторами мембраны постсинаптических нейронов и передавать информацию, они называются нейротрансмиттерами. Обычно такие рецепторы являются лигандуправляемыми каналами и очень быстро открываются в ответ на пресинаптический сигнал. Так, спустя всего несколько миллисекунд после прихода сигнала на постсинаптическую терминаль нейротрансмиттер, связываясь с рецептором постсинаптической мембраны, регулирует ионный поток (открывая или закрывая канал). К классу быстродействующих медиаторов принадлежат глутамат и гамма-аминомасляная кислота.
Медленные модуляторы
Медиаторы, ответственные за информационную модуляцию, действуют медленнее по сравнению с нейротрансмитеррами (рис.1). Для того чтобы изменить мембранный потенциал постсинаптического нейрона, модуляторам требуется время порядка сотен миллисекунд или нескольких секунд. Функция этих медиаторов заключается в модуляции (медленном изменении) действия быстродействующих медиаторов. Действие нейромодуляторов по сравнению с нейротрансмиттерами может длиться долгое время (но при этом оно слабое), поскольку запускаемые ими каскадные реакции могут протекать в течение нескольких дней. К классу нейромодуляторов принадлежат: моноамины (норэпинефрин, дофамин и серотонин), ацетилхолин и нейропептиды.
Модуляторные системы
В мозге существует четыре модуляторные системы: холинергическая, норадренергическая, серотонинергическая и дофаминергическая. Модуляторные системы характеризуются общими признаками:
- берут начало от относительно небольших отделов ствола мозга и базальных отделов переднего мозга;
- проецируются на ретикулярное ядро таламуса;
- обильно иннервируют кору, базальные ганглии, мозжечок и центры ствола мозга.

Cледует обратить внимание на то, что кластеры нейронов организованы на всем протяжении от спинного мозга до базальных отделов конечного мозга. Проекции от этих сравнительно немногочисленных нейронных образований иннервируют весь мозг и все периферические мышцы, органы и железы.
Холинергические и моноаминергические нейроны мозга формируют глобальные сети. Хотя такие нейроны происходят из достаточно локальных областей мозга, они занимают отделы спинного мозга, ствола мозга и базальных отделов конечного мозга (рис. 2) и иннервируют кору и подкорковые структуры, моделируя информационные процессы. Нейроны работают как единая сеть, генерируя широко распространяющиеся паттерны активности совместно в фазах сна и бодрствования, настроения (радость и печаль), когнитивных состояний внимания и медитации.
Холинергические нейроны сконцентрированно локализуются на уровне ядер среднего мозга, ядер базальных отделов переднего мозга и базальных ганглиев. Из септальной области холинергические нейроны проецируются в гиппокамп. Нейроны базальных отделов переднего мозга посылают проекции в ретикулярное ядро таламуса, орбитофронтальную и другие области коры. Считается, что холинергическая система играет ключевую роль в регуляции общей возбудимости и формировании памяти (активируя гиппокампальную систему). Ацетилхолин рассматривается как основной регулятор мощности гиппокампального тета-ритма, при этом предполагается, что причиной болезни Альцгеймера может являться гибель нейронов, продуцирующих ацетилхолин. Именно этим обусловлено использование препаратов, увеличивающих уровень продукции ацетилхолина мозгом (ингибиторы ацетилхолинэстеразы) в лечении нарушений памяти у пациентов пожилого возраста (при болезни Альцгеймера).
Большинство норадренергических нейронов локализуется на уровне ствола мозга в области голубого пятна. Нейроны голубого пятна проецируются в лобные области коры, через бета1-рецепторы, регулируя настроение. Проекции, для которых характерно преобладание альфа2-рецепторов, участвуют в модуляции процессов, связанных с вниманием и рабочей памятью. Норадренергические нейроны также имеют связи с лимбической корой (регуляция эмоций), мозжечком (регуляция моторных действий), кардиоваскулярными центрами среднего мозга (регуляция кровяного давления). Считается, что депрессия может быть обусловлена снижением продукции норадреналина и, наоборот, ее увеличение (относительно нормы) может приводить к мании.
Серотонинергические нейроны в основном локализуются в ядрах шва ствола мозга и иннервируют лобные отделы коры и структуры лимбической системы, обеспечивая регуляцию настроения и эмоций (тревожность и эмоции при боли). Серотонинергические нейроны ядер шва, иннервируя базальные ганглии, участвуют в модуляции движений, а также навязчивых состояний и действий. Проецируясь в гипоталамус, нейроны, продуцирующие серотонин, регулируют аппетит и пищевое поведение. Дополнительно, благодаря связям с центрами сна на уровне ствола мозга, модулируются состояния сна и бодрствования.
В дофаминергической системе мозга выделяют мезолимбический, мезокортикальный и нигростриарный проводящие пути. Дофаминергические нейроны черной субстанции в составе нигростриарного пути проецируются в базальные ганглии, которые играют определяющую роль в процессах выбора действий. Мезолимбический дофаминергический проводящий путь соединяет вентральную область покрышки ствола мозга с прилежащим ядром, которое, входя в состав базальных ганглиев, принимает участие в обеспечении работы лимбической системы и регулирует эмоциональное поведение — ощущение удовольствия, сильное чувство эйфории при употреблении наркотиков. Проекции мезокортикального дофаминергического пути, берущие начало от вентральной области покрышки ствола мозга, достигают дорсолатеральной префронтальной коры и поясной извилины (лимбической коры). Результатом фармакологических исследований, 1 доводившихся в течение 30 лет, явилось создание препаратов, способных изменять концентрацию дофамина в мозге человека. Было показано, что препараты, приводящие к увеличению концентрации дофамина, вызывают появление положительных психотических симптомов. Тогда как действие препаратов, снижающих концентрацию дофамина, проявляется в снижении выраженности этих симптомов. В частности, употребление веществ, характеризующихся стимулирующим действием (как, например, амфетамин или кокаин, увеличивающие уровень продукции дофамина), может вызвать параноидальный психоз, сравнимый с шизофренией. С другой стороны, все известные антипсихотические вещества (т.е. редуцирующие психотические симптомы) блокируют рецепторы дофамина. В частности, блокируются О2-рецепторы дофамина. Именно на этих представлениях и основана принимаемая многими специалистами теория психозов, известная как дофаминовая теория шизофрении.