Электроэнцефалография (ЭЭГ) — метод исследования головного мозга, основанный на регистрации его электрических потенциалов.
Понятие «электроэнцефалография» дуалистическое, объединяет в себе, по сути, два термина: методологию и методику.
Электроэнцефалография — раздел электрофизиологии, изучающий механизмы генерации суммарной биоэлектрической активности головного мозга, а также нейрофизиологигеская методика регистрации биоэлектрической активности головного мозга с конвекситальной поверхности головы.
Существуют и другие методы регистрации суммарной биоэлектрической активности головного мозга: электрокортикография — регистрация биоэлектрической активности непосредственно с коры, электросубкортикография — регистрация активности глубоких (подкорковых) структур мозга и др. Эти методы в подавляющем большинстве случаев используются в клинических целях для оценки локализации очага поражения головного мозга.
Многочисленные исследования показывают, что электрические потенциалы отдельных нейронов головного мозга связаны тесной и достаточно точной количественной зависимостью с информационными процессами.
История электроэнцефалографии
➥ Основная статья: История электроэнцефалографии
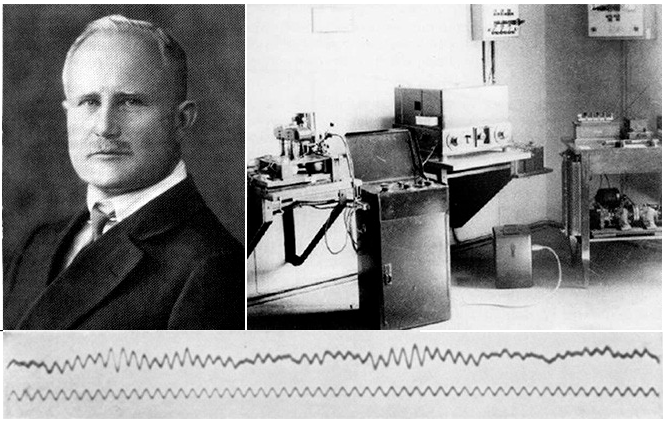
Первая публикация о наличии токов в центральной нервной системе была сделана Du Bois Reymond в 1849 г. В 1875 г. данные о наличии спонтанной и вызванной электрической активности в мозге собаки были получены независимо R.Caton в Англии и В.Я. Данилевским в России. Исследования отечественных нейрофизиологов на протяжении конца XIX и начала XX века внесли существенный вклад в разработку основ электроэнцефалографии. В.Я. Данилевский не только показал возможность регистрации электрической активности мозга, но и подчеркивал ее тесную связь с нейрофизиологическими процессами. В 1912 г. П.Ю. Кауфман выявил связь электрических потенциалов мозга с «внутренней деятельностью мозга» и их зависимость от изменения метаболизма мозга, воздействия внешних раздражений, наркоза и эпилептического припадка. Подробное описание электрических потенциалов мозга собаки с определением их основных параметров было дано в 1913 и 1925 гг. В.В.Правдич-Неминским.
Австрийский психиатр Ганс Бергер в 1928 г. впервые осуществил регистрацию электрических потенциалов головного мозга у человека, используя скальповые игольчатые электроды12. В его же работах были описаны основные ритмы ЭЭГ и их изменения при функциональных пробах и патологических изменениях в мозге. Большое влияние на развитие метода оказали публикации G.Walter3 о значении ЭЭГ в диагностике опухолей мозга, а также работы F.Gibbs, E.Gibbs, W.G.Lennox4, F.Gibbs, E.Gibbs (1952, 1964), давшие подробную электроэнцефалографическую семиотику эпилепсии.
В последующие годы работы исследователей были посвящены не только феноменологии электроэнцефалографии при различных заболеваниях и состояниях мозга, но и изучению механизмов генерации электрической активности. Существенный вклад в эту область внесен работами E.D.Adrian, B.Metthews5, G.Walter (1950), В.С.Русинова6, В.Е.Майорчик7, Н.П.Бехтеревой8, Л .А. Новиковой (1962), H.Jasper9. Большое значение для понимания природы электрических колебаний головного мозга имели исследования нейрофизиологии отдельных нейронов с помощью метода микроэлектродов, выявившие те структурные субъединицы и механизмы, из которых слагается суммарная ЭЭГ 1011.
Важнейшее значение для понимания механизмов генерации нормальной ЭЭГ и использования ее в качестве инструмента функциональной диагностики имели исследования неспецифических систем ретикулярной формации и лимбического комплекса — структур, определяющих уровень функциональной активности мозга121314.
Электроэнцефалографическая семиотика
Согласно определению, данному английским философом Джоном Локком (1632—1704), «семиотика — это учение о знаках». Название восходит к греч. σημειωτικός — «знак, признак». Специфический «язык», система специальных терминов, используемых для описания сути исследуемых явлений, входят в состав любой самостоятельной науки и учебной дисциплины. Единая система знаков неизбежно необходима для общения специалистов «на одном языке». Изучаемым предметом для ЭЭГ является биоэлектрическая активность головного мозга, что, собственно, и определяет смысловую нагрузку электроэнцефалографического «языка».
Таким образом, под электроэнцефалографической семиотикой следует понимать систему специальных терминов («знаков»), описывающих, классифицирующих и передающих информацию о параметрах суммарной биоэлектрической активности головного мозга.
Электроэнцефалографическая семиотика может быть представлена как иерархическая система. Ее основу составляет совокупность так называемых базовых терминов и понятий. По аналогии с известной каждому евклидовой геометрией, базовые ЭЭГ термины можно условно обозначить как систему аксиом. Аксиома (греч. ἀξίωμα — утверждение, положение) — это постулат, исходное положение, принимаемое в рамках какой-либо науки истинным без требования доказательства, то есть априорно. Аксиомы — краеугольные камни фундамента при построении любой науки. На фундаменте аксиом строятся доказательства всех последующих положений, которые, в свою очередь, называются теоремами.
К «ЭЭГ-аксиомам» следует, например, отнести такие понятия, как «отведение», «монтаж», «активность», «ритм», «паттерн», «синхронизация», «десинхронизация», «артефакт». Эти понятия бездоказательно принимаются как исходные постулаты электроэнцефалографии. На основе этих исходных понятий, строятся интерпретация и осмысление результатов исследований, доказываются электроэнцефалографические теоремы.
Классификация ЭЭГ
Следует различать области применения методики регистрации биоэлектрической активности головного мозга, поскольку у разных разделов нейронаук свои задачи.
- Клиническая ЭЭГ
- эпилептологическая
- неэпилептологическая
- ЭЭГ критических состояний
- физиологическая (психофизиологигеская) ЭЭГ
Предметом исследований ЭЭГ в клинике является биоэлектрическая активность головного мозга при заболеваниях (поражениях) головного мозга. Такие исследования могут быть определены как клиническая ЭЭГ. Клиническая ЭЭГ, в свою очередь, уже достаточно давно перестала быть однородной. Можно выделить, по крайней мере, три относительно самостоятельных направления: эпилептологическое, неэпилептологическое и ЭЭГ критических состояний (ЭЭГ-исследования, выполняемые в отделении реанимации при острой церебральной недостаточности).
Выполняются исследования в психологии и психофизиологии, в которых предметом рассмотрения выступает нейродинамика мозга здорового человека. Данный вид ЭЭГ можно определить как физиологическая (психофизиологигеская) ЭЭГ. Понимание специфических задач каждого из направлений и соотнесение их с методическими возможностями ЭЭГ позволяют эффективно выполнять исследования и получать глубокое удовлетворение от полученных результатов.
Физиология ЭЭГ
Головной мозг — это иерархически организованная система из нескольких десятков структурных образований с различным уровнем биоэлектрической активности. Электроэнцефалография отражает, в первую очередь, активность коры мозга, а также многих других структурно-функциональных образований. Для понимания физиологии ЭЭГ следует последовательно проследить, как электрофизиологические свойства отдельного нейрона формируют электроэнцефалограмму, а далее — оценить вклад различных структур мозга в суммарную активность.
Недостатки и ограничения электроэнцефалографии
Следует всегда понимать границы возможностей методики. Регистрация ЭЭГ позволяет оценить состояние системы генерации суммарной биоэлектрической активности головного мозга. «Работоспособность» механизмов биоэлектрогенеза выступает надежным нейрофизиологическим коррелятом функционального состояния центральной нервной системы (ЦНС). ЭЭГ, как всякая функциональная методика, не может локализовать структурные поражения. ЭЭГ не может по параметрам нейродинамики инвариантно констатировать нарушения психических функций и оценивать их тяжесть. Амплитудно-частотные параметры ЭЭГ слабо коррелируют с протеканием психических функций.
Преимущества метода
В отличии от ЭЭГ, такие нейровизуализационные методы, как «функциональный нейроимиджинг» — ПЭТ и СПЭКТ в большом числе случаев оказываются недостаточно чувствительными и специфичными, особенно на начальных стадиях заболеваний головного мозга или при преобладании метаболических нарушений над структурно-морфологическими как при некоторых формах склерозирующих и некротизирующих энцефалитов, дегенеративных и динамических ишемических нарушениях.
Регистрация ЭЭГ
➥ Основная статья: Регистрация ЭЭГ
Footnotes
- Berger H. Ueber das Elektroenkephalogramm des Menschen / Arch. Psychiat. Nervenkr., 1929, b. 87, s. 527-570.
- Berger.H. Ueber das Elektroenkephalogramm des Menschen I Arch. Psychiat. Nervenkr., 1932, b. 98. s. 231-254.
- Walter W.G. The localisation of cerebral tumors by electroencephalography I Lancet, 1936, v. 231, p. 305-308.
- Gibbs E, Gibbs E., Lennox W.G. Epilepsy: a paroxismal cerebral dysrhythmia / Brain, 1937, v. 60, p. 377-388.
- Adrian E.D., Matthews B.H.C. Interpretations of potential waves in cortex. J.Phisiol., 1934, p. 440-471.
- Русинов B.C. Некоторые вопросы теории электроэнцефалограммы. В кн.: Физиология. Л., 1954, с. 235—256.
- Майорчик В.Е. Электрофизиологический анализ функциональных свойств коры больших полушарий в зоне патологического очага. Физиол. журн. СССР, 1957, т. 3, с. 193-201.
- Бехтерева Н.П. Биопотенциалы больших полушарий головного мозга при супратенториальных опухолях. Л. 1960.
- Jasper H. Functional properties of the thalamic reticular system. In: The brain mechanisms and consciousness. A symposium. Oxford, 1954, p. 371-401.
- Костюк П.Г., Шаповалов А.И. Электрофизиология нейрона. В кн.: Современные проблемы электрофизиологических исследований нервной системы. М.: Медицина, 1964, с. 31-50.
- Eccles J.C. The Physiology of synapses. — Berlin-Gottingen-Heidelberg: Springer-Verlag, 1964.
- Анохин П.К. Нейрофизиологические основы электрической активности коры головного мозга. В кн.: Основные вопросы электрофизиологии центральной нервной системы. АН УССР, 1962, с. 132—163.
- Jasper H. Diffuse projection systems. The integrative action of the thalamic reticular system I Electroencephalogr. and Clin. Neurophysiol., 1949, v. 1, p. 405—419.
- Magoun H.W. The waking brain. Charles C. Tomas Publisher, Springfield, 111., USA, 1958.