Медленные волны сна — ЭЭГ-активность частотой от 0,3 до 1 Гц, доминирующая в ЭЭГ на стадиях глубокого сна. Данный тип активности генерируется благодаря кооперированным механизмам рекуррентных корковых цепей и связан с периодическими флуктуациями состояний покоя и бодрствования.
Up- и down-состояния
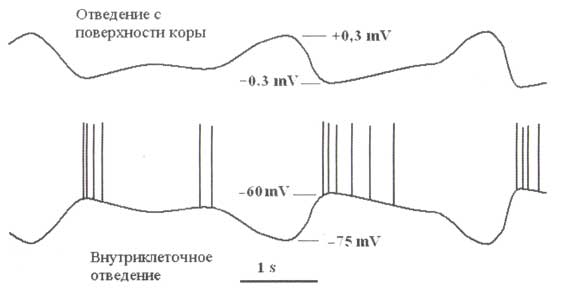
А. Медленные колебания локального внеклеточного потенциала. Б. Данные внутриклеточной регистрации. Обратите внимание, что отрицательные изменения внеклеточного потенциала совпадают с увеличением импульсной активности нейронов — up-состояние, в то время как положительные изменения внеклеточного потенциала связаны с подавлением нейронных разрядов — down-состояние. Обратите внимание также, что медленные колебания мембранного потенциала нейрона обратны по отношению к локальным полевым потенциалам. Адаптировано из Haider et al. (2006).
Медленные колебания — доминирующий паттерн во время медленноволнового сна и некоторых форм анастезии. Частота медленных колебаний — в пределах диапазона 0,3— 1 Гц. Активность в этой полосе частот также присутствует во время других состояний, таких как состояние с открытыми глазами, закрытыми глазами и во время решения различных задач. Однако вопрос, остается ли механизм возникновения этой активности во время бодрствования тем же, что и во время сна, еще требует ответа.
Наличие медленных колебаний после обширных таламических повреждений указывает, что медленные волны генерируются в пределах коры. Медленная волна представляет последовательность up- и down-состояний корковой нейронной сети. Up-состояние характеризуется активацией (увеличением частоты разрядов) корковых нейронов, связанной с деполяризацией этих клеток (рис. 1). Активность в этом состоянии постоянна в течение приблизительно половины секунды из-за четкого баланса между возбуждением и торможением в пределах текущей нейронной сети. Down-состояние характеризуется подавлением импульсной активности нейронов вследствие гиперполяризации корковых нейронов. Детальные нейронные механизмы генерации медленных волн остаются неизвестными. Один из гипотетических механизмов (Баженов и другие, 2002) состоит в том, что случайная суммация маленьких возбуждающих постсинаптических потенциалов в down-состоянии может активизировать постоянный ток натрия и деполяризовать мембрану выше порога генерации электрических спайков (разрядов).
Хотя медленные волны генерируются внутрикорковыми механизмами, периодические изменения между up- и down-состояниями упорядочивают таламические электрические паттерны (типа дельта-ритмов и сонных веретен) посредством кортико-таламических путей. Вследствие их синхронизирующего влияния на нейронную активность в пределах коры головного мозга и во взаимодействии с таламическими и гиппокампальными кругами, медленные колебания, как предполагается, лежат в основе консолидации памяти в течение сна.
Транскраниальная индукция медленных волн
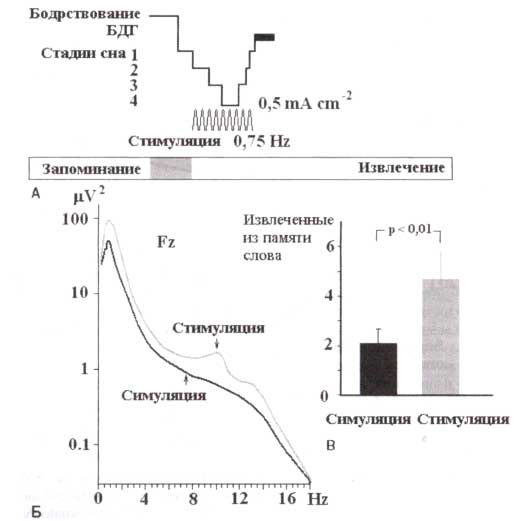
А. Временная диаграмма эксперимента. Обозначено время запоминания и извлечения информации из памяти, интервалы стимуляции, период выключения света (горизонтальная серая полоса) и сон, представленный гипнограммой. Б. Выполнение задач на декларативную память при условии стимуляций во время ночного сна и при условии ложных стимуляций. Эффективность выполнения выражается в виде разницы между числом правильно названных слов и общим числом слов, предложенных для запоминания. В. Диаграммы усредненной мощности спектров ЭЭГ в течение периодов с электрическими стимуляциями (серый столбик) и в периодов с ложными стимуляциями (черный столбик) в среднелобном отведении. Заштрихованные области указывают диапазоны частот медленных колебаний (0,5-1 Гц) и медленной лобной веретенообразной активности (верхний ряд, 8-12 Гц), адаптировано из Marshal et al. (2006). Обозначения: БДГ — фаза быстрых движений глаз.
Неясно, имеют ли медленные корковые потенциалы и их внеклеточный эквивалент какое-либо физиологическое значение сами по себе. Попытка проверить эту гипотезу была недавно предпринята применением внеклеточных медленных переменных токов1. Током воздействовали на молодых здоровых добровольцев через стимулирующие электроды, прикладываемые с двух сторон в лобно-латеральных положениях (участки анодной, положительной поляризации) и на мастоиды (рис. 2 вверху). Транскраниально применялись колебания тока в частоте 0,75 Гц с максимальной плотностью тика 0,5 мА/см2. Стимуляция начиналась через 4 минуты после того, как субъекты вступали во вторую стадию сна NREM (впервые после засыпания), во время, когда сон должен прогрессировать в стадию с медленными волнами. Субъекты были проверены дважды — в состоянии стимуляции и в состоянии ложной стимуляции. В каждом условии вечером перед сном субъекты разучивали различную информацию. Качество запоминания проверялось следующим утром. Декларативная память измерялась запоминанием ассоциативных пар (pained-associate leamingtask). Предъявлялся список из 46 пар слов, необходимо было правильно воспроизвести не менее 60 % (при немедленном предъявлении первого слова из пары (указывающего слова) была поставлена задача назвать второе слово). При проверке воспроизведения утром после сна указывающие слова показывались снова, и испытуемые должны были вспомнить соответствующее ответное слово.Показано, что стимулирование медленными токами с помощью транскраниального воздействия в течение раннего ночного сна увеличивает сохранение воспоминаний в памяти. Стимуляция вызвала увеличение продолжительности следующего промежутка медленноволнового сна и увеличение корковых медленноволновых колебаний и медленной веретеннообразной активности в коре (рис. 2 внизу).