К первичным ответам (ПО) относят электрическую реакцию коры больших полушарий, которая возникает на залп импульсов, приходящий из специфических афферентов.
ПО можно вызвать либо применением адекватных стимулов (свет, звук и т. д.) или электрическим раздражением рецепторов, чувствительных нервов, релейных ядер или проводящих путей того или иного анализатора, ПО, как правило, регистрируются в проекционных зонах анализаторов — в областях окончаний афферентов из специфических ядер таламуса.
Огромный экспериментальный материал, посвященный анализу ПО, еще в достаточной степени противоречив. В связи с этим возникают определенные трудности в трактовке генеза ПО и его функциональной роли. Поэтому мы вначале рассмотрим общие, более принятые взгляды на происхождение различных фаз ПО, а затем коснемся тех противоречий, которые возникли в последние годы в связи с более углубленным анализом их механизмов.
При регистрации суммарной активности с поверхности коры электродом площадью в 1 мм² в толще коры под электродом находится приблизительно 50 000 нейронов и около 100 000 афферентных и эфферентных волокон, не считая огромного числа глиальных элементов. Афферентный залп импульсов, приходящий в этот и соседний с электродом участки коры, вызывает сложную пространственную и временную динамику синаптических процессов, отражающихся на поверхности коры в виде различных фаз первичного ответа.
Обычно для ПО в коре существует фокус максимальной активности (ФМА), где амплитуда ПО максимальна. ФМА обычно находится передвижением электрода по поверхности изучаемой зоны коры. Как правило, используется униполярный способ отведения, Когда один электрод (индифферентный) помещается на костях черепа, а другой (активный) — на исследуемом участке коры.
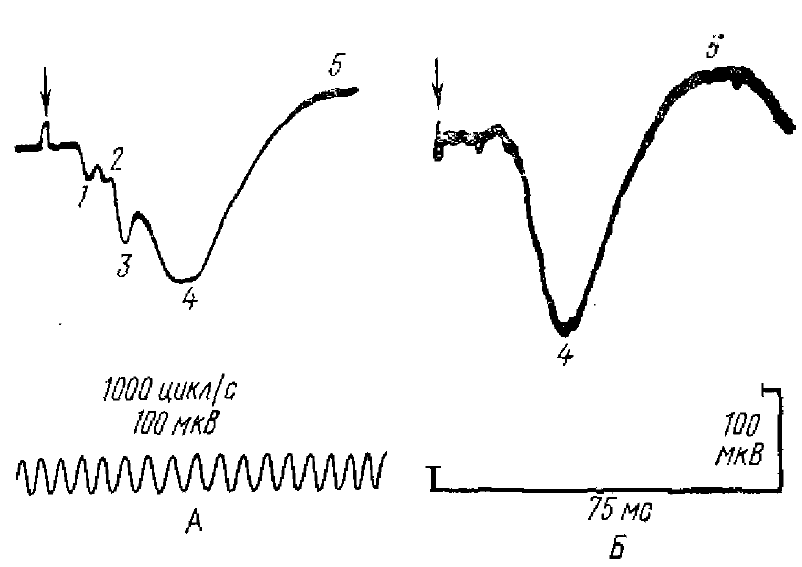
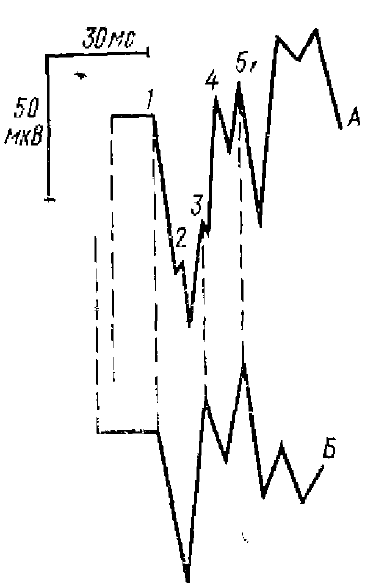
1, 2, 3 — быстрые колебания,4 — положительная и 5 — отрицательная фазы первичного ответа. Стрелкой показан момент раздражения
Учитывая сложную организацию нейронной сети коры, можно было бы ожидать и сложного характера суммарных колебаний потенциала, регистрируемого от ее поверхности. Однако форма коркового ПО крайне проста: он, как правило, состоит из двух основных колебаний (или фаз) — положительного и следующего за ним отрицательного (рис. 1). Простота формы ПО в значительной степени определяется особенностями строения коры и афферентных ее связей, а также компактностью и мощностью афферентного залпа импульсов, поступающих от специфических афферентов. С другой стороны, как мы увидим далее, за простотой формы, ПО кроется сложный характер нервных процессов, лежащих в основе генеза двух его основных фаз.
В различных корковых зонах анализаторов форма ПО варьирует, фазы ПО имеют различные временные характеристики, варьируют латентные периоды ПО, число, и выраженность быстрых волн, приходящихся в основном на положительную фазу ответа (см. рис. 1). Разная форма ПО определяется также различиями в функциональной организации тех или иных проекционных зон анализаторов, а также функциональным состоянием коры. Так, под наркозом, когда спонтанная активность коры подавлена, ПО наиболее хорошо выражены и становятся более стабильными по форме, а у бодрого, хронически оперированного животного форма и величина ПО значительно варьируют, появляются поздние компоненты ПО.
Латентный период ПО зависит от способа и интенсивности стимуляции, а также от числа синаптических переключений в специфическом афферентном пути. В значительной степени латентный период ПО изменяется в связи с состоянием сна или бодрствования, во время ориентировочной реакции или в процессе выработки временной связи, что часто определяется процессами проведения афферентного залпа на уровне релейных ядер таламуса. Однако для определенной проекционной зоны коры, при одной и той же форме и силе стимуляции и стабильных условий эксперимента, латентный период ПО, как правило, почти не меняется.
Происхождение различных фаз ПО
Прежде всего рассмотрим быстрые колебания, которые обычно наблюдаются на положительной фазе ПО. Так, у кошки быстрые колебания хорошо выражены у ПО зрительной коры и хуже у слуховой и соматосенсорной коры. Однако Рабин (1970) показал, что и в соматической области коры кошки при стимуляции передаточного таламического ядра число быстрых колебаний у ненаркотизированного животного может достигать пяти—семи, а у наркотизированного нембуталом двух—четырех.
Зрительный ПО содержит три быстрых волны (см. рис. 1). О происхождении этих колебаний существуют самые различные мнения. Ряд исследователей полагал, что все три колебания зрительного ПО являются отражением возбуждения трех групп зрительных афферентных волокон радиации (Chang, Kaada, 1950).
Чанг (Chang, 1952) выдвинул гипотезу, что эти колебания связаны с передачей в кору из сетчатки информации о цвете. Волокна, обладающие наибольшей скоростью проведения, несут «красную» информацию, средняя группа — «зеленую» и самые медленные — «голубую». Однако последующие исследования поставили под сомнение эту гипотезу. Был выдвинут ряд других соображений относительно природы этих колебаний.
Опыты со стрихнинизацией коры, локальным ее охлаждением и нагреванием, удалением коры и регистрацией активности от зрительной радиации и т. д, привели исследователей к выводу (Ройтбак, 1964; Bremer, Stoupel, 1959, и др.), что первое колебание связано с приходом в кору импульсации по волокнам радиации; второе и третье быстрые колебания являются отражением распространения импульсации по безмиэлиновым ветвлениям корковых волокон специфических афферентов.
Пресинаптическая природа быстрых колебаний была показана и для ПО соматосенсорной коры (Ройтбак, Бекая, 1960). Ройтбак (1961) считает, что охлаждение коры блокирует выделение медиатора в синаптических окончаниях, что приводит к блокаде положительной и отрицательной фаз ПО, постсинаптических по своей природе, в то время как быстрые колебания остаются. При локальном отравлении коры кокаином или нембуталом начальные быстрые колебания остаются также без изменений, в то время как положительная и отрицательная фазы ПО резко изменяются.
Существуют и другие точки зрения на быстрые колебания. Так, первое и второе колебания связывают с активностью афферентных волокон, третье — с возбуждением корковых нейронов или с возбуждением афферентных волокон связывают только первое колебание, в то время как второе и третье колебания с постсинаптическим возбуждением корковых нейронов (Purpura, Grundfest, 1956, и др.).
Постсинаптическую природу этих колебаний гипотетически объясняют разрядом сомы и аксонов последовательно разряжающейся цепочки нейронов в пункте регистрации (Bishop, Clare, 1952, 1953) или повторным разрядом пирамидных клеток.
Рабин (1970), Дуринян, Рабин (1971) считают, что они отражают повторный разряд пирамидных нейронов различных слоев коры, на которых оканчиваются таламокортикальные проекции, а более поздние из них — вторичную активацию нейронов за счет внутрикорковых связей.
Таким образом, тюка нет единой точки зрения на природу быстрых колебаний ПО, хотя очевидно, что решение этого вопроса представляет существенный интерес для выяснения генеза фаз ПО в связи с тем, что эти колебания безусловно отражают в той или иной степени функциональную организацию поступающего к коре афферентного залпа импульсов.
Положительный потенциал (фаза) ПО
Как правило, связывают с постсинаптическим возбуждением клеточных элементов III и IV слоев коры, возникающим под действием залпа афферентных импульсов. Так, при постепенном погружении отводящего электрода в глубь коры наблюдается вначале уменьшение положительной фазы ПО, а затем наступает инверсия ее знака на отрицательный с максимумом амплитуды в районе IV слоя коры. При экстра- и внутриклеточной регистрации от нейронов III—IV слоев коры многими авторами получены прямые доказательства, что глубинная отрицательная фаза ПО есть алгебраическая cyммa деполяризационных потенциалов. Обычно глубинный отрицательный потенциал является зеркальным отражением положительного колебания на поверхности коры. Показано также, что во время этого глубинного потенциала возникают спайковые разряды нейронов III—IV слоев коры.
Внутриклеточные исследования (Amassian et al., 1955) на соматосенсорной коре показали, что во время поверхностно-положительной фазы ПО в нейронах регистрируются ВПСП, на вершине которых возникают спайковые разряды. При слабых раздражениях глубинная негативность может, видимо, являться суммой ВПСП нейронов III—IV слоев коры, не приводящих к спайковому разряду нейронов. Интересные данные в этом отношении получил Сторожук (1970). Им было показано, что пришедший в кору афферентный залп одновременно вызывает деполяризацию большого числа нейронов. Лишь небольшое количество нейронов (например, спонтанно активных нейронов) реагируют генерацией импульсов. У подавляющего же большинства нейронов эта синаптическая бомбардировка не снижает величины мембранного потенциала до критического уровня, хотя амплитуда их ВПСП значительна. Он считает, что вклад в генез положительной фазы ПО таких не отвечающих спайком нейронов является наиболее весомым. Надо также полагать, что глубинный отрицательный потенциал является отражением не только ВПСП базальных дендритов и тел пирамидных нейронов III—IV слоя коры, но и нейронов, имеющих короткие аксоны и синаптические связи с афферентными волокнами. Так как большинство пирамидных нейронов посылает свои дендриты к поверхности коры, то в результате образующегося физического диполя с поверхности коры регистрируется положительный потенциал. Амплитуда поверхностно-позитивного потенциала может быть меньше глубинной отрицательности, так как не все нейроны посылают свои отростки на поверхность коры.
Итак, положительный потенциал ПО, по мнению исследователей, является дипольным отражением местного возбуждения пирамидных нейронов III—IV слоев коры. Однако в последние годы многие исследователи пришли к выводу, что только теорией физического диполя нельзя объяснить генез положительной фазы ПО. Появились данные, что инверсия положительной фазы обнаруживается не во всех опытах, наблюдается зависимость инверсии от вида наркоза и т. д. Ряд исследователей начинает связывать положительную фазу ПО со специализированной гиперполяризационной реакцией элементов поверхностных слоев коры (Аладжалова, Коштоянц, 1957; Гусельников, Супин, 1968; Кулланда, 1968; Батуев, Пирогов, 1969, 1971, и др.).
Так, «дипольная» теория положительной фазы ПО подразумевает, что глубинный отрицательный потенциал должен иметь равный (или меньший) латентный период, чем поверхностный положительный потенциал ПО. Данные, полученные К. М. Кулландой (1968, 1970), показали отсутствие «зеркальности» между поверхностным и глубинным ответами и опережение во времени положительным потенциалом отрицательного. Им было показано, что глубинный отрицательный потенциал имел даже больший латентный период (на 2,5—3,5 мс), чем поверхностный положительный. Таким образом, как отмечает Кулланда, «следствие возникает раньше причины, которая должна его обусловливать». Кроме того, инверсия положительного потенциала наступала в его опытах на глубине 1200—1400 мкм— на уровне V слоя коры — места расположения больших пирамид.
Эти и другие данные, полученные К. М. Кулландой, привели его к выводу, что генез положительной фазы ПО целиком связан с постсинаптическими процессами на дендритах коры. Он полагает, что положительная фаза ПО является отражением ТПСП апикальных дендритов пирамидных нейронов V слоя коры (Кулланда, 1968).
На участие в происхождении поверхностно-положительной фазы ПО гиперполяризующих синапсов указывал Гарсиа Рамос (Garsia Ramos, 1964). Пурпура и Грундфест (Purpura, Grundfest, 1956) также полагают, что фазы ПО отражают суммацию де- и гиперполяризационных ПСП, генерируемых в элементах глубоких слоев коры и апикальных дендритах. В нашей лаборатории (Супин, 1966; Гусельников, Супин, 1968) также были получены данные, что позитивная фаза ПО зрительной коры кролика связана с активной гиперполяризацией поверхностных структур коры и не является пассивным физическим отражением глубинно-негативной фазы. Используя парные стимулы, можно получить полную блокаду поверхностно-позитивной фазы при сохранении глубинно-негативной.
Специальный анализ генеза ПО в соматосенсорной и зрительной областях коры кошки был проведен А. С. Батуевым (1968), А. С. Батуевым и А. А. Пироговым (1970). Исследование инверсии положительной фазы ПО соматосенсорной коры показало, что глубинная негативность, как и в опытах К. М. Кулланды, возникала позже поверхностной позитивности на 1,6—2,5 мс и ни в одном из опытов не было их абсолютного совпадения. Специальный нейрохимический анализ ПО на световые раздражения у кошек, предпринятый А, И. Батуевым и А. А. Пироговым, показал, что ПО коры имеет сложную гетерохимическую природу, обе его фазы реализуются преимущественно в поверхностных слоях и отражают активацию как возбуждающих, так и тормозных механизмов.
Серков (1970), Василевский (1971) и др. также не нашли точного совпадения отрицательности в глубине коры с положительной фазой на поверхности. Сложный, не отвечающий «классическим» представлениям характер инверсии ПО зрительной коры кролика описан Полянским (1963).
Таким образом, накопившиеся в последние годы данные заставляют с новых позиций подойти к генезу положительной фазы ПО.
Надо полагать, что положительная фаза ПО является комплексным явлением, отражающим как сложное взаимодействие синаптических процессов возбуждения и торможения на различных уровнях коры, так и дипольного момента. Однако пока трудно представить, каким образом специфическая афферентная импульсация первоначально может активировать апикальные дендриты корковых нейронов.
Разные взгляды на генез положительной фазы ПО, возможно, объясняются тем, что данные, получаемые рядом исследователей, не совсем сопоставимы в связи с разными объектами исследования, условиями эксперимента И различиями в характере применяемых стимулов.
Отрицательная фаза ПО
По мнению большинства исследователей, является отражением возбуждения нейронных элементов поверхностных слоев коры. Если это положение почти не вызывает сомнения, то вопрос о том, каким образом возбуждаются поверхностные слои коры, является еще дискуссионным.
Отрицательная фаза ПО причинно связана с разрядами корковых нейронов. Об этом свидетельствуют многочисленные данные. Так, отрицательная фаза ПО не возникает в том случае, если нейроны в ответ на периферическое раздражение не разряжаются импульсами.
При слабых внешних раздражениях отрицательная фаза ПО не возникает, так как такое раздражение вызывает лишь местное возбуждение нейронов IV и III слоев коры, без спайковой активности. С поверхности коры в этом случае, как мы уже говорили, регистрируется лишь положительная фаза ПО. Не возникает поверхностно-отрицательная фаза ПО и при глубоком наркозе или аноксии, устраняющих разряды корковых нейронов на вершине глубинного отрицательного потенциала (соответствующего положительному потенциалу, ПО) (Li et al., 1956). При усилении разрядов корковых нейронов усиливается и отрицательная фаза ПО (стрихнинное отравление, усиление периферического раздражения и т. д.).
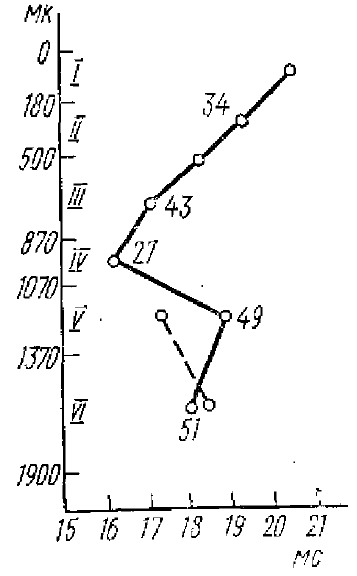
Цифры на кривой — количество зарегистрированных нейронов
Наиболее распространенная точка зрения на генез отрицательной фазы ПО связана с представлением о синаптическом возбуждении верхушечных дендритов через коллатерали, отходящие от аксонов возбужденных нейронов III и IV слоев коры. После прихода афферентного залпа импульсов к Ill и IV слоям, благодаря этим коллатералям, возбуждение распространяется к элементам I и II слоев коры. По данным Паттона и Амассиана (Patton, Amassian, I960), латентный период разряда нейронов больше в верхних слоях коры и наиболее короткий в III—IV слоях (рис. 2). Однако надо заметить, что такие зависимости рядом исследователей не были найдены (Сторожук, 1968; Серков, 1970 и др.).
На возникновение отрицательной фазы ПО за счет потенциалов, генерируемых нервными элементами поверхностных слоев коры, указывает факт ее инверсии. При углублении электрода в кору из средних и глубоких ее слоев во время отрицательного потенциала на поверхности регистрируется позитивный потенциал. В пользу связи отрицательной фазы ПО с верхними элементами коры говорит также факт избирательного подавления отрицательной фазы путем поверхностной термокоагуляции коры.
Другая точка зрения связана с представлением о том, что отрицательная фаза ПО является результатом распространения возбуждения от сомы пирамидных нейронов III и IV слоев вверх по их апикальным дендритам в I слой коры. Это представление подкреплялось тем фактом, что при постепенной проходке отводящим электродом толщи коры наблюдается постепенное уменьшение латентного периода отрицательного потенциала (или постепенное уменьшение длительности положительного потенциала). Затем на определенном уровне коры положительный потенциал уступает место отрицательному. Кроме того, данные по распределению отрицательного поверхностного потенциала ПО в вертикальном направлении по коре показали градуальность его изменений, свидетельствующую о непрерывном его распространении. Однако вопрос об антидромном распространении возбуждения по дендритам корковых нейронов, как мы уже говорили ранее, остается пока открытым.
Таким образом, какой бы точки зрения на генез отрицательной фазы ПО не придерживаться, обе они связаны с тем представлением, что положительная и отрицательная фаза ПО причинно связаны между собой.
Однако в последние годы получены новые данные, которые заставили по-иному трактовать генез отрицательной фазы ПО.
Другая точка зрения на генез отрицательной фазы ПО была выдвинута П. К. Анохиным (1969) на основании исследований Ф. А. Ата-Мурадовой (1961, 1963) и Пурпуры (Purpura, 1961). По концепции П. К, Анохина, положительная фаза ПО связана с приходом возбуждения от специфических афферентов, в то время как отрицательная фаза — от неспецифических афферентов. Таким образом, обе фазы имеют различную природу и причинно между собой не связаны.
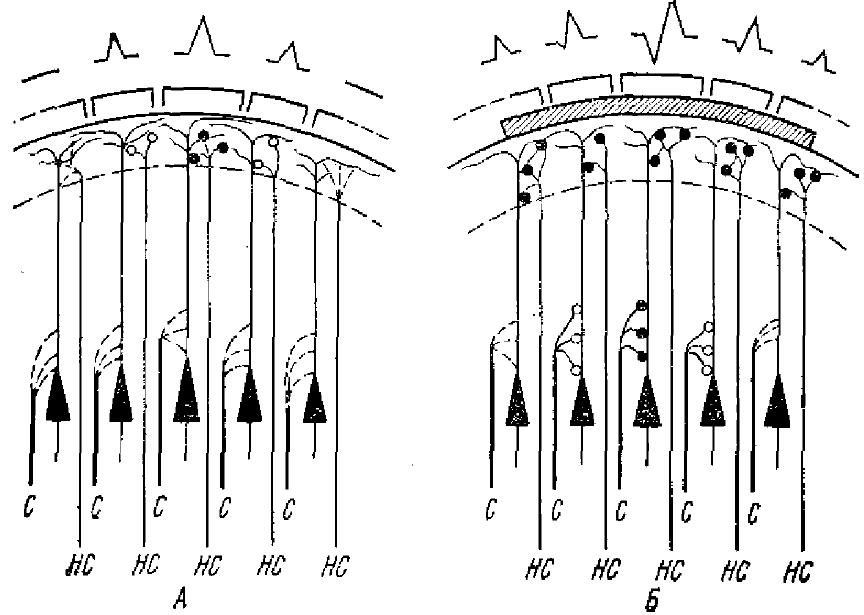
А — видно созревание отдельных синаптических образований в плексиформном слое (обозначены кружочками); в третьем и четвертом слоях синаптических образований не имеется; Б — видно прогрессирующее созревание синапсов плексиформного слоя, соответствующее распространению отрицательного потенциала по коре больших полушарий. Вместе с тем видно начало созревания аксо-соматических синапсов, что соответствует появлению положительной фазы отрицательного потенциала. Характер вызванных потенциалов и их распространение по коре схематически показано вверху; С — специфические афферентные пути, НС — неспецифические афферентные пути
Какие основные экспериментальные данные легли в основу такой концепции. Известно, что у млекопитающих в процессе постнатального онтогенеза наблюдается гетерохронное созревание на корковых нейронах специфических и специфических синаптических окончаний. Первыми созревают аксо-дендритические синапсы от неспецифических афферентов и только спустя определенное время формируются аксосоматические синаптические связи специфических афферентов (рис. 3). С другой стороны, в лаборатории П. К. Анохина, Ф. Л. Ата-Мурадовой было показано, что в начальные, стадии постнатального онтогенеза раздражение седалищного нерва вызывает в проекционной области соматосенсорной коры кролика локальный вызванный потенциал отрицательной полярности, он регистрируется в ограниченном пункте коры. С ростом животного зона его отведения расширяется. Морфологическими исследованиями (Ата-Мурадова, Чернышевская, 1961) было выяснено, что он формируется в коре возбуждением, приходящим по волокнам неспецифического происхождения, которые восходят из белого вещества к апикальным дендритам пирамидных клеток. Связь неспецифических афферентных волокон с молекулярным верхним слоем коры была описана Лоренте де Но и Фултоном еще в 1943 г. и другими исследователями.
Самые разные воздействия на этот отрицательный потенциал (разный наркоз, нагревание поверхности коры и т. д.) показали, что он имеет совершенно самостоятельный генез и, видимо, причинно не связан с положительной фазой ПО, которая появляется только к 7—8-му дню постнатального онтогенеза в связи с началом функционального созревания специфической таламо-кортикальной проекционной системы.
В связи с этим любое регистрируемое на поверхности коры гомогенное электрическое колебание может объясняться множественным характером восходящих влияний в кору больших полушарий (Анохин, 1968). Иными словами, можно ожидать, например, что отрицательная фаза ПО отражает не одну, а несколько посылок из подкорковых структур афферентных залпов импульсов разного генеза и функционального значения.
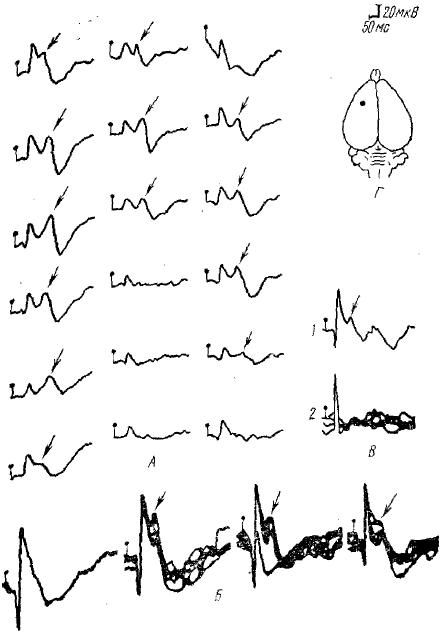
А — различные варианты соотношения первичного или вторичного (указано косой стрелкой) отрицательных компонентов у четырехдневного кролика при отведении из одного и того же пункта коры. У кроликов старшего возраста обе негативности слиты. Однако, во время нембуталового наркоза начинает проявляться вторичная негативность, а первичная уменьшается; Б — различные формы объединения отрицательных компонентов в отрицательную фазу на 15 день; В — изолированный первичный отрицательный компонент (1). зарегистрированный а одном ИЗ пунктов коры и на расстоянии 3 мм от него — вторичный ответ с двухкомпонентной отрицательной фазой (2); Г — схема мозга: черная точка — пункт отведения потенциалов. Начало раздражения везде обозначено вертикальной черточкой с кружком
Исследования, проведенные Ф. А. Ата-Мурадовой (1967), Ф. А. Ата-Мурадовой и Л. М. Чуппиной (1970), действительно показали двойственную природу отрицательной фазы ПО соматосенсорной коры кролика. Онтогенетический анализ отрицательной фазы ПО выявил, что в процессе постнатального онтогенеза она складывается из двух отрицательных компонентов, один из которых наблюдается уже у новорожденных, а второй созревает позднее. Авторы отмечают, что оба компонента весьма отчетливо регистрируются до 15—20 дня раздельно в связи с дисперсией формирующих их восходящих возбуждений, но по мере созревания коры они начинают сливаться друг с другом (рис. 4).
Применяя различные дозы нембуталового наркоза, а также местное действие (аппликация) растворов стрихнина и нембутала, Л. М, Чуппипа (1970), Ф. А. Ата-Мурадова и Л. М. Чуппина (1971) нашли, что отрицательная фаза соматосенсорного ПО взрослого кролика состоит из двух отрицательных компонентов: первичного и вторичного. Эти компоненты неодинаково генерализованы по коре, обладают различной чувствительностью к применяемым фармакологическим препаратам и т. д. Это дало авторам возможность сделать заключение о том, что они отражают на поверхности коры активность двух различных восходящих систем с разными подкорковым генезом и синаптическими организациями в коре. Пространственное перекрытие и синхронная активация этих систем в зрелой коре приводит к слиянию двух раздельных отрицательных колебаний в общую отрицательную фазу ПО. Авторы считают, что первичный отрицательный компонент в онтогенезе созревает первым и имеет локальную зону проявления в коре. Вторичный — созревает позднее, имеет более широкую зону проекции в коре, как и вторичный ответ Форбса (см. лекцию 7). Разное действие нембутала и стрихнина на эти ответы можно, видимо, объяснить разной специфичностью химии постсинаптических мембран соответствующих систем синапсов.
Однако надо отметить, что не все исследователи разделяют такую точку зрения на генез отрицательной фазы ПО.
Д. А. Фарбер (1969) пришла к выводу, что связи коры больших полушарий со специфической подкорковой системой переключения функционируют с момента рождения. Начально отрицательный характер ВП коры новорожденного животного, по ее мнению, связан с особенностью приема афферентной сигнализации по специфическим афферентным путям и с незрелостью воспринимающего нервного аппарата коры больших полушарий. Она показала, что при афферентных раздражениях и прямом электрическом раздражении специфических ядер таламуса регистрируются однотипные ответы. Это дало ей основание полагать, что регистрируемый в раннем постнатальном периоде начально-негативный ВП обусловлен приходом возбуждения по специфической афферентной системе. Его негативность она связывает с приходом возбуждения по специфическим афферентам к поверхностным слоям коры (а не к III—IV слоям).
Послойная регистрация в коре этого ПО на прямое раздражение специфических зрительных релейных ядер таламуса действительно показала уменьшение и инверсию негативного потенциала уже на глубине 100— 200 мкм. У взрослых животных основная масса специфических афферентов формируется уже на уровне нейронов Ш—IV слоев коры и с момента прозревания животного негативная волна начинает регистрироваться на уровне этих слоев, отражаясь на поверхности коры в виде позитивной фазы ответа. У новорожденных крольчат не наблюдается импульсной реакции нейронов на вспышку света, соответствующей по времени ВП, она наблюдается только с появлением первичной позитивности ВП. Поэтому Д. А. Фарбер (1968) считает, что начально-негативный ВП имеет аксодендритное происхождение (ПСП апикальных дендритов), а первичная позитивность, вернее, соответствующая ей глубокая негативность — отражает активацию сомы нейронов III—IV слоя. Что касается новорожденных детей, то локальные ПО, регистрируемые Фарбер в проекционной зоне коры на световую стимуляцию, по форме не отличались от ответов взрослого. Они содержат все основные компоненты ПО, в том числе и начальную позитивность.
Таким образом, по данным Д. А. Фарбер и других исследователей (Scherrer, Occonomos, 1954; Дзидзишвили, Джавришвили, 1961), начально-негативная фаза ПО у новорожденного животного и отрицательная фаза ПО у взрослых имеют разное происхождение, хотя обе они определяются залпом импульсов из специфических афферентов.
В раннем онтогенезе это происходит за счет прямой активации специфическими афферентами поверхностных слоев коры, хотя пути такой активации и гипотетичны, у взрослых животных активируются в основном III и IV слои коры с последующей активацией верхушечных дендритов через коллатерали корковых нейронов.
Шеррер и Экономо (1954) предположили, что отрицательное колебание в раннем онтогенезе по мере созревания животного эволюционирует в положительное колебание. Они полагают, что отрицательный знак ответа в раннем онтогенезе объясняется поверхностным расположением тел клеток III—IV слоя, по мере созревания поверхностного нейропиля клетки III—IV слоев постепенно отодвигаются в глубь коры, что и приводит к начальной позитивности первой фазы ПО на определенной стадии онтогенеза. Эту точку зрения поддержали Дзидзишвили и Джавришвили (1961). Однако, исходя из этого, трудно объяснить, что в процессе онтогенеза наблюдается увеличение амплитуды отрицательного колебания (Ата-Мурадова, 1963), в то время как должен был иметь место обратный процесс в связи с удалением внутрь коры структур, его генерирующих.
Необходимо также отметить, что не во всех случаях у новорожденных животных ПО состоит из чисто отрицательного колебания. Так, Пурпура (Purpura, 1961) зарегистрировал ПО в виде положительно-отрицательного комплекса. Положительный потенциал ПО был выражен только при прямой электрической стимуляции таламо-кортикальной радиации. В этом случае в кору поступает более синхронный залп афферентных импульсов, чем в случае адекватных раздражений.
В опытах с прямым раздражением специфических ядер таламуса позитивная фаза ПО у новорожденного кролика при большой силе тока была зарегистрирована также Д. А. Фарбер (1969). Это показывает, что уже на ранних стадиях онтогенеза можно зарегистрировать обычный двухфазный ПО. В этом случае можно полагать, что слабая выраженность (или отсутствие) положительной фазы ПО на ранних стадиях онтогенеза объясняется лишь большой дисперсией афферентных импульсов в незрелых, еще медленно проводящих афферентных путях.
Таким образом, мы пришли к выводу, что к настоящему времени еще нет единого мнения о генезе различных фаз ПО. И если на первых этапах изучения генеза ПО кажущаяся элементарность его двухфазового колебания в какой-то мере обнадеживала исследователей, то по мере углубления анализа его механизмов все больше становится ясным, что это сложная реакция.
Действительно, ПО является многокомпонентной реакцией, особенно при регистрации его в условиях хронического эксперимента и при применении адекватных стимулов (Fourment, 1958; Альтман, Марусева, I960, и Др.). В этом можно убедиться на примере ПО зрительной коры.
В хиазме выявляется пять положительных компонентов (1—5) с латентными периодами соответственно: 12—13; 19— 20; 25—26. 30—31 и 36—38 мс
В нашей лаборатории многокомпонентность ПО зрительной коры кролика была показана А. Я, Суминым (1961) и Л. М. Мухаметовым (1962). Так, А. Я. Суини нашел, что ПО состоит из двух двухфазных колебаний, наложенных друг на друга, и объяснил это наличием двух залпов импульсов из сетчатки. Л. М. Мухаметов, регистрируя одновременно суммарную активность зрительного перекреста и зрительной коры кролика на вспышку света, нашел несколько компонентов в хиазме, которым соответствовали различные фазы ПО (рис. 53). Многокомпонентность ПО и в этом случае можно было бы объяснить сложным залпом импульсов из сетчатки.
В работе на собаках Н. Н. Любимов (1964) нашел, что ПО на свет является суммарным ответом это, по его мнению, связано с проведением импульсации по трем специфическим каналам зрительного анализатора: по классическому зрительному пути, коллозальным волокнам из симметричной зрительной корковой зоны, а также в результате переключения на другую сторону на уровне переднего двухолмия.
Ф. А. Ата-Мурадова и Т. И. Белова (1971) также показали, что вызванный ответ зрительной коры кролика является по своей природе весьма сложным и каждая его фаза определяется возбуждением различной природы, Они нашли, что в процессе онтогенеза компоненты его созревают гетерохроний, что связано с гетерохронным созреванием восходящих к коре зрительных возбуждений (рис. 5).
Становится очевидным, что генез ПО прямо связан со структурой данной области мозга, с распределением и характером синаптических связей, функциональным состоянием данной области мозга, различной системой афферентных проекций в кору, со сложной мозаикой пространственного и временного течения процессов возбуждения и торможения на разных уровнях анализатора и особенно в высшем его отделе — коре больших полушарий. Надо также полагать, что ПО различных проекционных зон коры имеют также существенные различия, определяемые спецификой структуры, связей и функциональной организацией разных уровней того или иного анализатора. Раскрыть эти особенности ПО — насущная задача будущих исследовaтелей.
Иррадиация ПО по коре больших полушарий
Локален ли ПО в проекционной зоне коры или он распространяется по коре в горизонтальном направлении? На этот счет существуют два диаметрально противоположных мнения.
Один исследователи полагают, что ПО распространяются за пределы первичных воспринимающих областей коры (Clare, Bishop, 1954; Артемьев, 1962; Ткаченко, Шульга, 1964; Ткаченко, 1963; Шульга, 1965, 1966; Коган, Ткаченко, 1969, и многие другие).
Этой точке зрения противопоставляется другая, в которой ряд исследователей считает, что ПО является локальной реакцией и не распространяется по коре больших полушарии. Потенциалы, имеющие короткий латентный период и регистрируемые за пределами проекционных зон анализаторов, эти исследователи относят к принципиально иным вызванным потенциалам — к категории ассоциативных ответов (Amassian, 1954; Buser et al., 1955, 1957, 1959, Moulders, 1962; Thompson et al., 1963, и многие другие). В этом случае предполагается, что широкая регистрация вызванных потенциалов за пределами проекционных областей на одно и то же раздражение связана исключительно с восходящей подкорково-корковой афферентацией, с афферентным потоком импульсов из ассоциативных ядер таламуса в ассоциативные области коры больших полушарий. Таким образом, кажущаяся генерализация по поверхности коры ПО является на самом деле отражением лишь вертикального распространения к разным зонам коры афферентных потоков из различных таламических образований, потоков разного генеза и функционального значения. Учитывая афферентные связи ассоциативных и неспецифических ядер таламуса, их проекционные отношения с корой, а также связи этих ядер с группой специфических ядер, возможности такой «вертикальной» активации коры крайне многообразны.
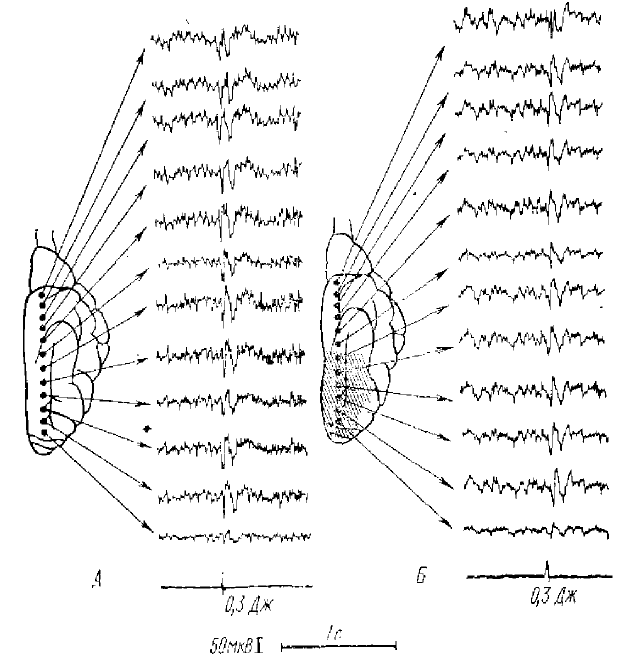
Регистрируется активность одновременно от 12 пунктов коры. Внизу — отметка вспышки, интенсивность в Дж
Однако, не отрицая этого положения, А. Б. Коган полагает, что существуют и электрические ответы в коре, которые являются отражением нервных процессов, развивающихся в первичных проекционных зонах анализатора на залп импульсов из специфических ядер таламуса (ПО) с последующей иррадиацией процессов по коре (рис. 6). Этот вывод подкрепляется многочисленными данными лаборатории Л. Б. Когана, которые показали, что в областях, примыкающих к проекционной зоне зрительного анализатора, а также в слуховой и соматосенсорной проекционных зонах можно зарегистрировать вызванные потенциалы, по форме сходные с ПО проекционной зрительной зоны. Отсюда, по мнению автора, потенциалы, регистрируемые вне проекционной области, в отличие от ПО являются отражением комплекса как восходящих, так и транскортикальных конвергенций.
Несколько по-иному к этому вопросу подошли И. Ю. Беленков и Ж. А. Белоусова (1970). Изучая коротколатентные ответы на зрительное раздражение в обширных зонах коры больших полушарий, они показали преимущественное значение «вертикальных» восходящих путей в широкой иррадиации зрительной импульсации. На основании опытов с удалением зрительной проекционной зоны (полей 17, 18, 19), с перерезкой мозолистого тела и отведением биопотенциалов по методу Лаплассиан они пришли к выводу, что ВП на свет в незрительных областях не связаны с возникновением ПО в зрительной коре и что они определяются импульсацией, поступающей в них по вертикальным таламо-кортикальным путям.
Об изолированном течении возбуждения в коре свидетельствуют и данные Маунткастла (Mountcastle, 1957). Активация колонки нейронов в соматосенсорной коре не распространяется на соседние колонки нейронов, которые могут возбудиться только при раздражении других рецепторов того же участка кожи. Известно также, что во время прихода афферентного залпа в кору вначале вовлекаются в активность много нейронов, по уже во время течения отрицательной фазы ответа они начинают тормозиться (Кондратьева, 1964; Полянский, 1966; Гусельников, Супин, 1968, и др.). Этот и ряд других фактов в какой-то мере трудно объяснить, если принять, что процессы, лежащие в основе ПО, иррадиируют по коре.
Механизмы изменчивости ПО коры
Как мы уже отмечали, ПО изменчив как по своей амплитуде, так и по выраженности фаз в зависимости от условий эксперимента и применяемых раздражений. Особенно изменчив он при хронических формах эксперимента и при самых различных дополнительных воздействиях на организм животного или человека. Прежде всего, характер ПО находится в прямой зависимости от процессов, разыгрывающихся в данный момент в нейронной сети изучаемого участка коры — от функционального состояния ц. н. с. Эти изменения функционального состояния могут быть вызваны чисто физическими или химическими факторами (давлением электродов, повреждением или подсыханием коры, изменением температуры коры, применением наркотиков и других физиологически активных веществ и т. д.). Изменения ПО прямо связаны с характером стимуляции, силой, длительностью и крутизной нарастания стимула, его спектральным составом (в случае световых раздражений), а также с биологической его значимостью. Кроме того, характер ПО зависит от регулирующих влияний подкорковых центров и, в первую очередь, ретикулярной формации (РФ) ствола мозга, оказывающих прямое влияние на возбудимость корковых элементов и на релейные станции, передающие специфическую импульсацию к коре. Существенному изменению подвергается ПО в процессе ориентировочной и условнорефлекторной деятельности. ЦНС контролирует уже первое звено афферентной части рефлекторной дуги, рецепторные образования — место формирования афферентного залпа к коре больших полушарий. Сейчас становится ясным, что взаимодействие различных анализаторов не ограничивается корковыми связями, — это взаимодействие может осуществляться уже на рецепторном уровне. Так, например, звуковые, болевые и другие виды раздражений могут оказывать влияние на возбудимость элементов сетчатки посредством эфферентных ее связей с РФ ствола, а также, возможно, и через центрифугальные волокна коры.
РФ оказывает влияние и на проведение афферентного залпа по специфическому пути. Известен факт снижения амплитуды ВП во всех звеньях специфических афферентных систем при ориентировочной реакции, т. е. при активации РФ она оказывает и облегчающее действие на передачу афферентного залпа импульсов (при ее прямом раздражении или раздражении неспецифических ядер таламуса). Известно также, что регулирующее влияние РФ подчинено корковому контролю и, видимо, контролю лимбической системы, что значительно усложняет рефлекторную систему влияний на афферентный поток импульсов в специфических проводящих путях.
И наконец, важное значение имеют, видимо, экстраретикулярные пути контроля проведения импульсов в системе специфических проекции, а также прямые кортико-таламические связи в различных сенсорных системах, функциональное значение которых сейчас усиленно изучается. В физиологических экспериментах было выяснено, что эти влияния могут иметь тормозную или возбуждающую природу, а по данным других исследователей, они могут оказывать двойное действие — возбуждающее и тормозное. Интимные механизмы этих влияний еще не совсем ясны.
Таким образом, изменчивость ПО связана со сложной системой регуляции афферентного потока импульсов, имеющей, видимо, прямое отношение «к первичной фильтрации информации, уменьшению ее общего объема и дифференцированию функционально различных информационных потоков» (Р. М. Мещерский, I960).
Что касается изменчивости ПО при условнорефлекторной деятельности, то этому вопросу посвящено много работ, которые привели пока к противоречивым результатам. Одни исследователи полагают, что ПО увеличивается на тот раздражитель, который приобрел сигнальное значение. Другие полагают, что вообще нет никакой корреляции между изменениями ПО и образованием временной связи.
Ю. Г. Кратин (1967, 1971), изучая изменчивость ПО при различных формах условнорефлекторной деятельности, пришел к выводу, что качество сигнала — его значение как положительного или отрицательного раздражителя — не находит прямого отражения в ПО.
О трудностях использования ПО в работе на человеке для самых различных целей, в том числе и в области психологии, говорит в своей обзорной работе Уттал (Uttal, 1967). Можно полагать, что все «флюктуации» латентности, полярности, амплитуды и других параметров ПО связаны, прежде всего, с различной степенью деятельности общих механизмов активации и торможения, вызывающих сильные суммарные сдвиги функционального состояния коры и подкорковых структур мозга. Кроме того, использование ПО как электрографического показателя тонкой динамики условнорефлекторной деятельности вряд ли полностью оправдано, так как к настоящему времени нет четких данных о его генезе. На это обращает внимание и Куллаида (1971), говоря о том, что «эффективность дальнейшего использования ВП (в том числе и ПО) в качестве инструмента анализа закономерностей работы мозга находится в прямой зависимости от создания системы общих представлений о механизмах их генеза».
В последние годы намечается тенденция связывать так называемые поздние компоненты вызванных потенциалов с проведением возбуждения через неспецифические структуры среднего и промежуточного мозга (Иваницкий, 1971), а изменение их амплитуды ставить в зависимость от значимости раздражителя, его сигнального значения для организма, от уровня внимания испытуемого и т. д. (Пеймер, 1971; Chapman, 1966, и др.), для выяснения корково-подкорковых взаимодействий в функциональных системах различного биологического значения (Шумилина, 1971) и т. п.
Однако к этим поздним компонентам, следующим за ПО коры, разные авторы относят самый широкий спектр различных реакций: ВО, медленную волну, следующую за ПО, разряды последействия и т. д. Кроме того, во многих работах далеко не ясно, о каких реакциях конкретно идет речь, не ясен и генез этих потенциалов. Поэтому делать определенные заключения об отражении в «поздних компонентах» информационной деятельности мозга вряд ли было бы оправданно без глубокого и всестороннего анализа происхождения самих этих компонентов. Особенно это касается работ, проводимых на животных.
Информационное значение первичного ответа
Одной из неясных проблем, касающихся всех вызванных потенциалов, в том числе и ПО, является их функциональная значимость. До сих пор окончательно не ясно, какую информацию о сигнале отражают ПО.
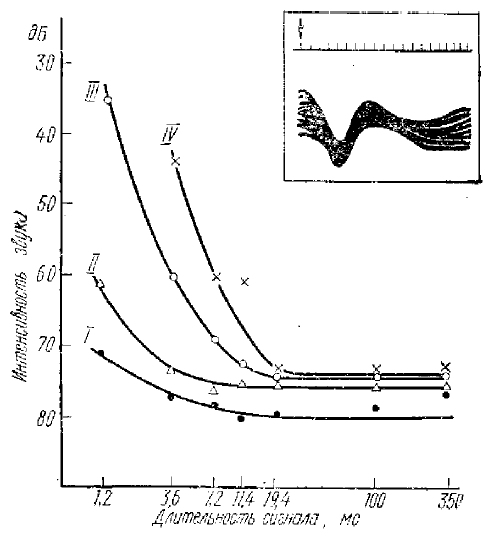
Параметр кривых — постоянные времени нарастания сигнала в мс: I — 0,2, II—0,6, III — 6,0, IV —20. Наверху справа — первичный ответ при надпороговой интенсивности (+3 дБ); отметка времени 7 мс, стрелка — начало раздражения
Вопрос об информационном значении ПО был поднят Гершуни (1962, 1965). Изучая зависимость между длительностью звукового сигнала и порогами возникновения ПО в слуховой области коры кошки, он нашел, что наблюдается закономерное понижение порогов при возрастании длительности сигнала, происходящее в некотором пределе времени (рис. 7). После достижения этого предела возрастание длительности раздражения не оказывает уже влияния на пороги и другие характеристики ПО. Это критическое время оказалось равным (в зависимости от крутизны нарастания амплитуды звукового сигнала) 7—20 мс. Отсюда им было высказано предположение, что ПО отражает работу специального механизма проекционных зон коры, который обеспечивает обнаружение и различение коротких звуков.
И. А. Пеймер (1971), исследуя ВП у человека, показал зависимость фаз ВП от свойств внешнего раздражителя, его сигнального значения, информации, содержащейся в стимуле, а также от формирования ответной моторной реакции. Однако ряд исследователей высказывают обоснованные сомнения в информационном значении ПО, что также имеет экспериментальные основания.
А. А. Василевский (1971) предположил, что расшифровка функционального значения ПО (как и других медленных электрических процессов мозга) и его связь с информационными характеристиками сигнала могут быть даны лишь в результате выяснения их корреляции с активностью отдельных нейронов. Известно, что амплитуда ПО зависит не только от силовых, но и от частотных параметров стимула. Он также показал, что у наркотизированных животных устойчивую амплитуду компонентов ПО можно наблюдать только при низких частотах стимуляции 10—15 кол/с. Однако в отличие от ПО вызванная импульсная активность отдельных групп нейронов отчетливо наблюдается вплоть до частот 30—100 кол/с. Из данных многих авторов известно, что при высоких частотах стимуляции в нейронах развивается устойчивый деполяризационный сдвиг мембранного потенциала и в этом случае достаточно совсем небольшой амплитуды ВПСП для генерации нейронного спайка. Суммарный ПО при таких высоких частотах не регистрируется, хотя «информация» о параметрах внешнего раздражителя воспринимается нейронами проекционной области коры.
Как мы указывали, ПО может явиться хорошим показателем общих изменений функционального состояния головного мозга. Однако, как показал Ю. Г. Кратин (1970, 1971), ПО не отражает таких изменений функционального состояния мозга, которые наступают в результате воздействия сигналов, несущих определенную информацию. Он указывает, что период времени для сбора информации, определяемый латентным периодом ПО, слишком мал. Эта информация, видимо, носит чисто ориентировочный характер, а «для полного анализа сигнала, после чего лишь может наступить адекватное изменение функционального состояния мозга и соответствующая биологическая реакция, требуется значительно большее время, особенно для тонкого различения раздражителей». Изучая изменения ПО слуховой коры кошки на звуки разного сигнального значения, он пришел к выводу о невозможности существенного отражения в ПО сигнального значения стимула. Качество сигнала не находит прямого отражения в ПО. Изменения ПО в этих случаях связаны, видимо, с ориентировочной реакцией. Надо полагать, что ПО отражает лишь реакцию коры на афферентный поток импульсов и не связан с процессами обработки информации о параметрах стимула (Швырков, 1969; Джон, 1967; Латаш и др., 1970).
С другой стороны, сам ПО в некотором отношении искусственный феномен, возникающий на раздражители, редко встречающиеся в естественных условиях обитания животного, где имеют место, как правило, медленно нарастающие по силе раздражения, биологически более значимые, но не приводящие к возникновению ПО. Известно (Забоева, 1966), что по мере увеличения постоянной времени нарастания сигнала уменьшается амплитуда всех фаз ПО и при постоянной времени порядка 300 мс ПО на звук полностью исчезает.
Таким образом, сложные формы нервной деятельности в большинстве своем связаны с дисперсным залпом импульсов в проводящих путях, со сложными процессами пространственно-временного анализа и синтеза сигнала в нейронной сети на всех уровнях анализатора, часто не имеющих четкого выражения в суммарных электрических процессах мозга. Однако, надо полагать, что вскрытие генеза ПО еще даст, хотя и в ограниченной сфере применения, ключ к ряду важных механизмов мозговой деятельности.
Необходимо отметить, что ПО были с успехом использованы для изучения локализации функций в корковых зонах анализаторов. При определении зоны регистрации в коре ПО на адекватную (или электрическую) стимуляцию рецепторной части анализатора была обнаружена весьма сложная организация пространственной проекции слухового, зрительного, обонятельного, соматосенсорного и интероцептивного анализаторов.
Так, например, слуховая область коры кошки по локализации ВП была разбита на 6 различных отделов (Woolsey, 1961). Были составлены также карты частотной локализации в слуховой коре собаки (Tunluri, 1950, 1960).
Многими исследователями проведено изучение сложной картины корковых зон организации соматосенсорного и двигательного анализаторов у животных и человека (Adrian, 1941; Marshal et al., 1941; Rose, Mountcastle, 1959, и многие другие) (рис. 8, 9).
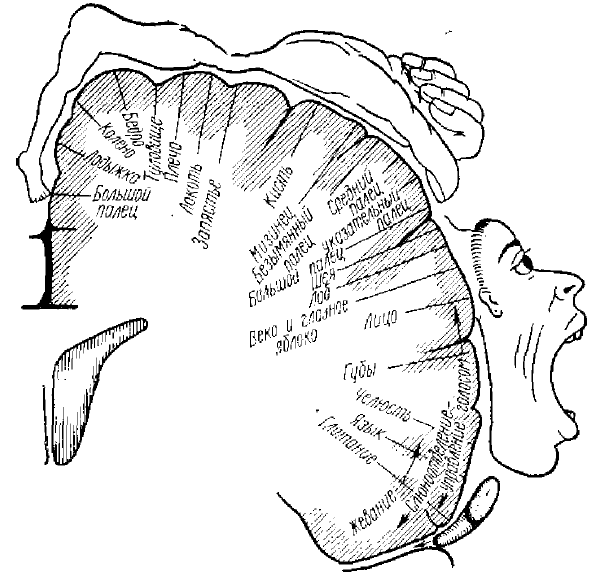
Части тела, изображенные вдоль центральной извилины на поперечном разрезе мозга, показывают размер протяженности соответствующих проекционных зон
Особенности организации центральных зон зрительного и других анализаторов с использованием ПО были изучены на кролике (Bishop, O’Leary, 1936; Thompson et al., 1950; Полянский, 1963 и мн. др.) и других видах животных: рыбах, амфибиях, рептилиях и птицах (Гусельников, 1965).
Представительство в коре висцеральных систем было подробно изучено В. Н. Черниговским и его сотрудниками (Черниговский и др., I960, 1962, 1970) (рис. 10), а также многими зарубежными исследователями.
Необходимо только отметить, что в связи со сложностью трактовки локальности ПО в настоящее время, наличия ВП в коре с латентным периодом, равным или даже меньшим, чем латентный период ПО и т. п., ряд данных по локализации функций в коре с использованием ПО, видимо, требует определенной коррекции и проверки. Надо полагать, что проекции в кору сенсорных систем значительно сложнее. Они включают в себя потоки первичной информации разного функционального значения, имеющие местом своего приложения отнюдь не узкую зону коры, очерченную как ядро анализатора.
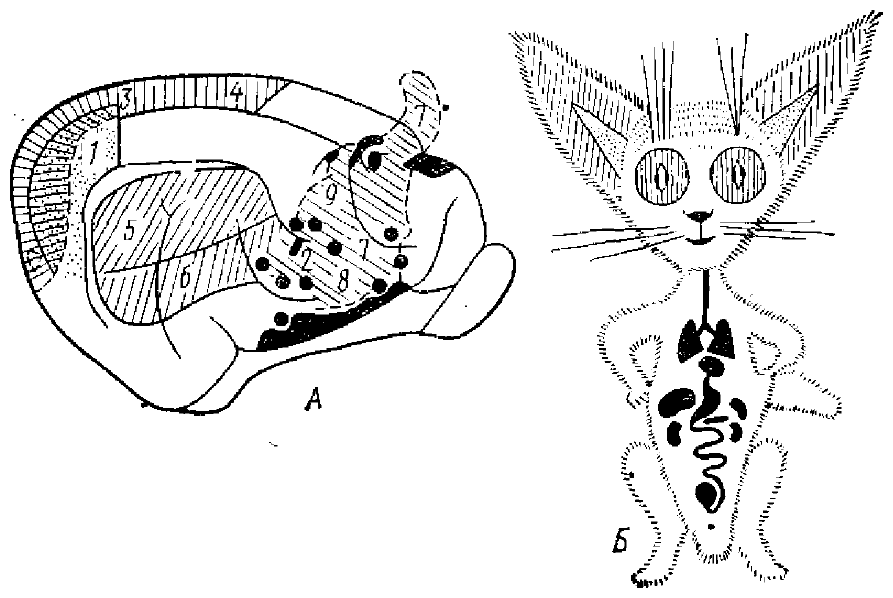
1 и 2 — первая и вторая вестибулярные зоны; 3 и 4 — первая и вторая зрительные зоны; 5 и 6 — первая и вторая слуховые зоны; 7, 8, 9 — первая, вторая и третья соматосенсорные зоны. На схемах корковых зон (А) черным выделены зоны представительства висцеральных систем, штриховкой — золы представительства соматосенсорных систем