Пик-волна является наиболее стабильной и специфической формой ритмики, характеризующей petit mal— эпилепсию, во время которой на поверхности коры мозга регистрируется широко генерализованная электрическая активность, представляющая собой повторение с постоянной частотой (3 в 1 с) сложного комплекса потенциалов, состоящего из отрицательных пика и волны длительностью 200—250 мс. Чрезвычайная устойчивость ритмики этого вида в эпилептической ЭЭГ привлекает внимание исследователей, несмотря на сравнительную редкость заболевания petit mal — эпилепсией.
Показано, что аналогичная активность может быть сравнительно легко получена в структурах, отличных от неокортекса человека и других млекопитающих, например, в древней коре лягушки и ящерицы, а также в переднем мозгу рыб (Servit et al., 1965, 1966, 1969). Наличие комплекса пик-волна у рыб показывает, что для генерации этой ритмической активности не обязательна корковая структура. Это говорит об общности главных принципов, лежащих в основе возникновения активности, для всего филогенетического ряда нервных структур и придает изучению механизмов ритмики типа пик-волна не только практический, но и теоретический интерес. Отсюда, можно также полагать, что и нейрофизиологический механизм этой активности может быть более прост по сравнению с другими видами ритмической активности.
В ходе изучения ритмики пик-волна выявилось три основных способа получения ее в эксперименте на животных. Первый при помощи ритмического электрического раздражения медиальных ядер таламуса (Ingvar, 1955; Pollen et al., 1963, и др.), второй фармакологическими воздействиями на мозг. Так, значительное число веществ — метразол, хлоралоза, хлорамбуцил, фторуксусная кислота, препарат S-335-47, алюминиевая паста и другие препараты — вызывают у теплокровных животных ритмику пик-волна. Третий способ провокации ритмики пик-волна связан с шоковой электрической стимуляцией коры. Однако он чаще применяется в работе на изолированной коре млекопитающих.
Мы в своих опытах (Петухов, 1968, Гусельников, Петухов, 1970, 1971), выполненных на кошках, использовали два первых способа. В случае ритмической стимуляции подкорковых структур (с частотой 3 в 1 с) мы получили активность типа пик-волна не способную к самоподдержанию и длящуюся только во время стимуляции. В дальнейшем активность этого вида мы будем называть вызванной. В качестве фармакологического агента, вызывающего ритмику пик-волна, мы использовали хлоралозу в смеси с барбитуратами. В этом случае ритмика, появляющаяся в определенных стадиях наркоза, характеризуется способностью к самоподдержанию, и поэтому в дальнейшем мы будем называть ее «спонтанной».

1-9-ЭЭГ, соответствующие точкам отведения, обозначенным на карте мозга (внизу). Отрицательному потенциалу соответствует отклонение луча вверх
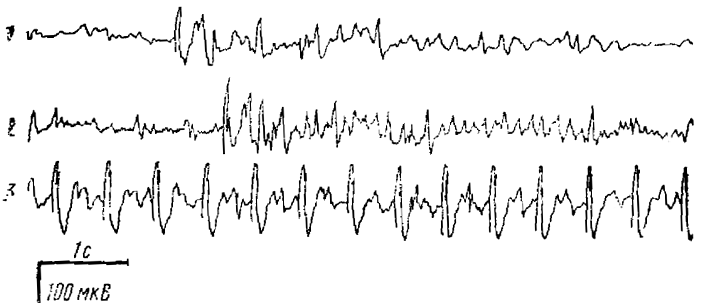
При раздражении интраламинарных ядер таламуса ритмическими электрическими импульсами с частотой около 3 имп/с в коре в ответ на каждый стимул возникает реакция, состоящая из двух основных компонентов; относительно короткого поверхностно-негативного колебания, по форме и длительности сходного с волной реакции вовлечения и поверхностно-негативной, но значительно более длительной — порядка 150—250 мс — волны, которая следует за этим колебанием (рис. 1). Такая форма реакции наблюдается только при указанной частоте стимуляции и не возникает ни при более частой стимуляции, ни при слишком редких таламических раздражениях. Последнее указывает на то, что этому типу ответов, так же как и реакции вовлечения, свойственна способность к рекрутированию при ритмической стимуляции.
По-видимому, интраламинарный таламус является не единственным местом, раздражением которого может быть вызвана такая активность. Аналогичные ответы типа пик-волна описал Вейр (Weir, 1964) при раздражении РФ ствола мозга.
Представляет интерес сопоставление медленных волн реакции типа пик-волна, получаемой при низкочастотном раздражении неспецифических структур таламуса, с медленной поверхностно-негативной волной, возникающей в коре, при достаточно сильном прямом ее раздражении. Мы уже указывали, что сильное электрическое раздражение поверхности коры может вызвать помимо начального коротколатентного «дендритного» ответа также и поверхностно-негативную волну, которая имеет ряд признаков, сближающих ее с негативной волной реакции типа пик-волна. Длительность негативной волны, возникающей при прямом раздражении коры (150—300 мс), практически совпадает с длительностью негативной волны реакции пик-волна. При регистрации от глубоких слоев коры можно наблюдать как для поверхностно-негативной волны на раздражение коры (Goldring et al., 1959), так и для волны реакции пик-волна (Pollen et al., 1964) характерную картину реверсии; при постепенном погружении регистрирующего электрода в глубину коры амплитуда волны уменьшается, доходит почти до нуля, а затем начинает вырастать волна противоположной (позитивной) полярности. Наконец, на сходство природы этих двух явлений указывает их чувствительность к фармакологическим агентам. Исследование импульсных разрядов нейронов коры мозга во время ритмической активности типа пик-волна, вызываемой низкочастотным (3 имп/с) раздражением интраламинарного таламуса, провели на моторной, соматосенсорной и ассоциативной коре мозга кошки Поллен и сотрудники (Pollen et al., 1964, 1969). Нейроны, отвечавшие на неспецифические раздражения, обнаруживали четкую корреляцию импульсной активности с поверхностной электрокортикограммой при появлении активности пик-волна. Во время более короткой (пикоподобной) поверхностно-негативной волны наблюдалось учащение импульсной активности нейронов. Медленная поверхностно-негативная волна (в глубине коры ей соответствует позитивная волна) сопровождается угнетением импульсной активности нейронов вплоть до полного торможения.
Эти результаты были подтверждены и уточнены с помощью внутриклеточной регистрации активности нейронов (Pollen, 1964, 1968). Было показано, что оба компонента комплекса являются постсинаптическими потенциалами нейронов коры. Начальная позитивность, которая может предшествовать негативному пику (при сильных раздражениях), является результатом деполяризации нейронов в глубоких и средних слоях коры (Pollen, 1968).
Пикоподобная негативная волна сопровождается ВПСП корковых нейронов, который имеет приблизительно такую же длительность (обычно до 25—40 мс и лишь в отдельных случаях — до 80 мс) и начинается практически одновременно с началом поверхностно-негативного пикоподобного колебания или на несколько миллисекунд позже. После этого начального ВПСП в подавляющем большинстве нейронов регистрируется длительный (обычно 120—180 мс, иногда до 280 мс) ТПСП, который имеет значительно больший латентный период, чем ВПСП и начинается через 19—30 мс после начала поверхностнонегативного пикоподобного колебания, почти одновременно с началом медленной поверхностно-негативной волны. Длительность ТПСП близка к длительности медленной поверхностно-негативной волны. ТПСП были регулярными и хорошо выраженными в тех случаях, когда в поверхностной ЭКоГ был хорошо выражен комплекс пик-волна.
Длительные ТПСП можно было зарегистрировать только при очень поверхностном барбитуровом наркозе, когда низкочастотное (3 имп/с) раздражение таламуса вызывало активность типа пик-волна. Углубление наркоза вело к подавлению медленных поверхностно-негативных волн и редукции ТПСП, тогда как начальные ВПСП и пикоподобные колебания сохранялись.
Все эти данные указывают на тесную связь пикоподобного колебания комплекса пик-волна с начальным ВПСП, а последующей медленной волны с ТПСП нейронов коры. Поверхностно-негативное пикоподобное колебание расценивается как аналог классических корковых потенциалов реакции вовлечения. Как и потенциалы реакции вовлечения, это колебание имеет негативную полярность при регистрации от поверхности коры и сопровождается деполяризационными колебаниями (ВПСП) нейронов более глубоких слоев. Поэтому, точно так же как и для волн реакции вовлечения, можно допустить, что пикоподобное колебание комплекса пик-волна генерируется деполяризующими синапсами, расположенными на поверхностных разветвлениях апикальных дендритов пирамидных нейронов коры, и вызывается непосредственно импульсами, приходящими в кору от раздражаемого таламического ядра. То, что комплекс пик-волна содержит компонент, аналогичный потенциалам реакции вовлечения, не является неожиданным, так как различие в условиях вызова активности типа пик-волна и реакции вовлечения заключается только в некотором изменении частоты стимуляции тех же самых таламических ядер.
Медленная поверхностно-негативная волна комплекса пик-волна генерируется, очевидно, тормозными гиперполяризующими синапсами: она сопровождается ТПСП и блокадой импульсной активности корковых нейронов. Но, поскольку волна имеет на поверхности коры негативную полярность, она может генерироваться гиперполяризующими синапсами, расположенными не вблизи поверхности коры, а в более глубоких слоях. Создаваемый синапсами ток, направленный из клетки во внеклеточную среду, вызовет позитивный сдвиг потенциала глубоких слоев коры, по отношению к которым поверхность коры окажется более негативной. Медленная позитивная волна в глубоких слоях коры, соответствующая по времени поверхностно-негативной медленной волне, действительно регистрируется (Pollen et al., 1964), что подтверждает правильность приведенного объяснения.
Исследование чувствительности поверхностно-негативной волны к различным фармакологическим агентам и поляризации коры (Pollen et al,, 1964) позволяет несколько уточнить механизмы ее генерации. Было показано, что аппликация на поверхность коры блокатора синаптической активности — гамма-аминомасляной кислоты (ГАМК) — приводит к увеличению медленной поверхностно-негативной волны, тогда как пикоподобная волна угнеталась. Анодная и катодная поляризация коры также противоположным образом влияет на пикоподобную и на медленную поверхностно-негативную волну. Отсюда был сделан вывод, что гиперполяризующие синапсы, генерирующие медленные ТПСП при возникновении активности типа пик-волна, локализованы не только в глубоких, а во всех слоях коры до ее поверхности. Активность глубоко расположенных гиперполяризующих синапсов вызывает позитивную волну в глубине коры и негативную волну на поверхности, а активность поверхностно расположенных гиперполяризующих синапсов вызывает на поверхности позитивную волну. Позитивный и негативный потенциалы на поверхности коры складываются и взаимно погашаются, но, согласно гипотезе авторов, погашение является неполным: активность глубинных гиперполяризующих синапсов превалирует над активностью поверхностных, так что в результате на поверхности коры регистрируется негативная волна. Если же активность поверхностных синапсов заблокировать аппликацией ГАМК или катодной поляризацией, то частично замаскированная ранее активность глубинных синапсов выявляется полностью; следствием этого является увеличение поверхностно-негативной волны, что и наблюдалось в экспериментах.
О тормозной природе медленной волны комплекса свидетельствуют и наши данные (Гусельников, Петухов, 1968, 1970, 1971). Тормозные, нейроны, ответственные, за генерацию волны, расположены, видимо, на уровне коры, так как ответы пик-волна могут быть вызваны и в изолированной коре.
Ряд наблюдений привел Поллена и соавторов к выводу, что причиной, вызывающей развитие ТПСП на телах пирамидных нейронов коры, может быть включение механизма возвратного торможения, сходного в общих чертах с тем, который описан Андерсеном с сотрудниками для таламуса и некоторых других структур. Запуск системы возвратного торможения тем эффективнее, чем сильнее стимул подается на вход системы, что, видимо, связано с более высоким порогом возбуждения вставочного тормозного нейрона.
Согласно данным Джаспера и Поллена, активность типа пик-волна, наблюдаемая при ритмической стимуляции (3 в 1 с) неспецифического таламуса, приурочена в основном к ростральным корковым областям. Наши данные подтвердили это. При стимуляции самых различных ядер неспецифического таламуса активность типа пик- волна наблюдалась в соматосенсорной коре и ассоциативных областях коры и никогда не наблюдалась в затылочной коре. Некоторые детали топографии активности даны на рис. 1. Лучше всего волны выражены в ассоциативных ростральных областях коры, несколько хуже — в первичных проекционных областях, особенно в их медиальных отделах. Следует указать, что эти черты топографии активности сохранялись при стимуляции самых разнообразных ядер неспецифического таламуса, хотя степень выраженности волн по коре в целом зависела от места стимуляции. Так, наиболее четкие волны возникали при стимуляции ретикулярного и передневентрального ядер таламуса. Это, видимо, связано с тем, что при стимуляции других, более каудальных ядер неспецифического таламуса афферентный залп испытывает значительную дисперсию в ходе многочисленных синаптических переключений, характерных для неспецифического таламуса.
При стимуляции nucl. reticularis и nucl. ventralis anterior число переключении минимально, поскольку эти ядра проецируются непосредственно на новую кору, что определяет большую мощность и компактность приходящего в кору афферентного залпа, а следовательно, и более эффективный запуск системы возвратного торможения и генерацию волны. В связи с этим понятна лучшая выраженность волн в ассоциативных областях коры, где преобладают неспецифические афференты. В отличие от Поллена, который указывал на билатеральность активности типа пик-волна, мы наблюдали ее только при билатеральной стимуляции неспецифического таламуса. Унилатеральная стимуляция в большинстве случаев не сопровождается четкой активностью в противоположном полушарии.
Исходя из гипотезы о генерации медленной отрицательной волны системой внутрикоркового возвратного торможения, следует предположить, что стимуляция неспецифического таламуса должна быть лишь частным методическим приемом для получения в коре рассматриваемой активности. Действительно, ряд данных говорит за то, что медленные отрицательные потенциалы, по форме и длительности очень сходные с волной в комплексе пик-волна, развиваются в коре при самых разнообразных способах ее стимуляции. Кроме прямого раздражения коры длительный (до 400 мс) отрицательный потенциал возникает при стимуляции специфического восходящего пути (в частности, зрительного). В этом случае после первичного ответа возникает РРП из одной или нескольких отрицательных волн длительностью 200— 300 мс каждая.
➥ Более подробно: Первичные ответы коры больших полушарий
Мы получали в соматосенсорной коре кошек медленные отрицательные волны при ритмической стимуляции не только неспецифического таламуса, но и релейного соматического ядра — nucl. ventralis posterolateralis (рис. 2), а также хвостатого ядра (рис. 3). Однако пик, характерный для стимуляции неспецифического таламуса, отсутствовал. При стимуляции релейного соматического ядра каждый стимул сопровождался развитием медленной отрицательной волны, которой предшествовал слабо выраженный первичный ответ. Одиночные стимулы сопровождались длительным σ-послеразрядом. При стимуляции хвостатого ядра каждый стимул сопровождался развитием коротколатентного отрицательного пика и медленной отрицательной волной. Отрицательный потенциал обладал значительно меньшей латентностью, чем в комплексе пик-волна, развивающемся в коре при стимуляции неспецифического таламуса. Зависимость формы реакции от места стимуляции служит подтверждением того, что пик в комплексе пик-волна, видимо, действительно является аналогом пика реакции вовлечения, так как последний развивается в соматосенсорной коре лишь при стимуляции неспецифического таламуса; при стимуляции других ядер развиваются соответствующие им потенциалы (например, первичный ответ при стимуляции nucl. ventraiis posterolateralis).
Отведение от латерального края задней сигмовидной извилины. Раздражение отмечено артефактами

Поскольку развитие в коре комплекса пик-волна определяется только силой стимула, следовало предположить, что волна будет развиваться и в ответ на одиночный стимул. Действительно, одиночная стимуляция неспецифического и специфического таламуса, а также хвостатого ядра вызывала в коре развитие медленной отрицательной волны. При этом во всех трех случаях она являлась частью сложного комплекса, состоящего собственно из волны, предшествующей ей коротколатентной реакции и σ-послеразряда, аналогичного вызванному веретену. Если коротколатентная реакция определяется местом приложения стимула, то волна и σ-послеразряд во всех трех случаях одинаковы.
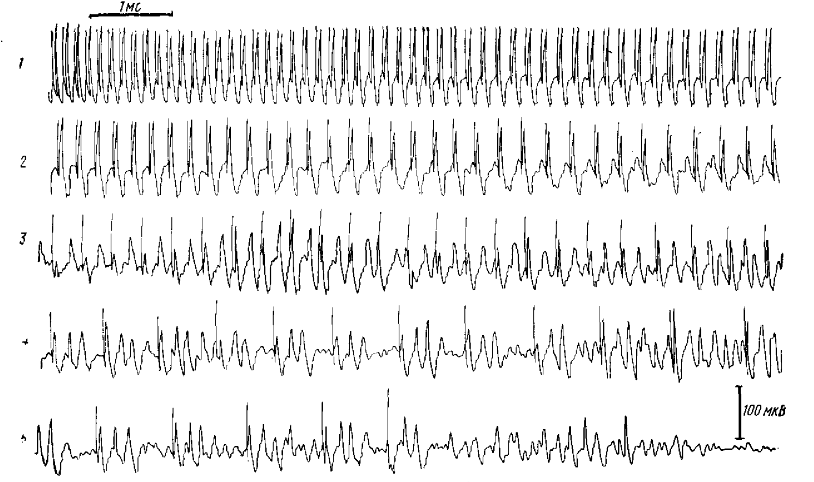
1 — частота стимуляции 8—9 Гц, 2 — 2—4 Гц, 3 — 3 Гц, 4 — 2 Гц, 5 — одиночный стимул. Каждый стимул отмечен артефактом.
Частоту стимуляции 3 в 1 с также не следует считать обязательной для появления активности пик-волна; она развивается и при других частотах стимуляции при условии, если интервал между двумя последовательными стимулами достаточен для развития волны. При этой частоте активность максимально приближается по форме к клинической картине petit mal — эпилепсии, поскольку сопровождающий волну послеразряд прерывается последующим стимулом. Следует отметить, что такое прерывание осуществляется на любой фазе развития комплекса: коротколатентная реакция — волна — σ-послеразряд. На рисунке 4 (1) видно, что благодаря этому на частотах стимуляции 8—9 Гц наблюдается типичная картина реакции вовлечения; на частоте 5 Гц (рис. 4, 2) видно начало волны, которая так и не успевает развиться полностью и обрывается. С дальнейшим урежением частоты стимуляции (рис. 4, 3, 4) наблюдается типичная активность пик-волна, и, наконец, на одиночный стимул (рис. 4, 5) развивается весь комплекс, включая ритмический послеразряд.
Таким образом, ни место, ни частота стимуляции не являются факторами, определяющими развитие в коре медленной отрицательной волны. Единственное определяющее условие — сила стимула. В этом отношении компоненты упомянутого комплекса отличаются по порогу. Наименьшим порогом обладает коротколатентная реакция, наибольшим — σ-послеразряд. Поэтому генерацию медленной отрицательной волны в коре в ответ на пришедший к ней сильный стимул надо рассматривать как закономерное явление. Следует отметить, что место стимуляции определяет степень генерализации этого потенциала в коре.
Степень генерализации определяется обширностью восходящих проекционных путей от стимулируемой структуры к коре. На основании данных о локальности медленной волны при прямой стимуляции коры следует считать, что генерация медленного отрицательного потенциала осуществляется в каждом отдельном участке коры самостоятельно. В связи с этим транскортикальная передача становится в интактной коре неэффективной уже на расстоянии 3—4 мм. Следовательно, внутрикорковая система, ответственная за генерацию медленной отрицательной волны, должна характеризоваться, во-первых, равномерным распределением по всей коре, во-вторых, чрезвычайной однородностью, поскольку результат ее активации в любом участке коры одинаков. Таким требованиям может отвечать лишь система, обладающая сравнительной однотипностью составляющих ее элементов и связей между ними. Структура механизма возвратного торможения должна, видимо, хорошо соответствовать этим требованиям. Дополнительные данные в пользу участия этой системы в генерации медленной отрицательной волны были получены нами в серии опытов по исследованию взаимодействия волны с вызванным потенциалом (Гусельников, Петухов, 1970).
Можно представить себе следующую схему генерации в коре всей последовательности потенциалов в ответ на пришедший в нее афферентный залп. Вначале возникает коротколатентная реакция; форма ее определяется входом, на который подана стимуляция. При прямой стимуляции коры это дендритный потенциал, при стимуляции специфического пути — первичный ответ, при стимуляции неспецифического таламуса — пик реакции вовлечения и, наконец, при стимуляции хвостатого ядра — быстрый отрицательный потенциал. Независимо от вида коротколатентной реакции результатом ее является активация корковых пирамидных нейронов. При достаточной силе возбуждения оно вызывает, во-первых, включение внутрикорковой системы возвратного торможения и, во-вторых, за счет корково-таламических связей включение таламической системы возвратного торможения. В результате в коре генерируется медленная отрицательная волна, а в таламусе «запускается» водитель σ-ритма, который, в свою очередь, по восходящим таламо-кортикальным связям вызывает генерацию σ-ритма в коре. После окончания волны нейроны в коре, подвергшиеся действию ТПСП, некоторое время сохраняют повышенную возбудимость и в этих условиях способны с большей вероятностью включаться в разнообразные активирующие процессы, в том числе и σ-ритм, имеющий подкорковый источник.
Основной особенностью вызванных волн, принципиально отличающей их от активности, наблюдаемой при petit mal — эпилепсии, является отсутствие у них способности к самоподдержанию, т. е. комплекс генерируется в пропорции один стимул — одна волна. Потенциально система возвратного торможения склонна к ответу серией ритмических колебаний. Не исключено, что для соматической коры пропорция один стимул — одна волна обусловливается наличием подкоркового водителя σ-ритма, который, захватывая облегченные после ТПСП нейроны, препятствует повторению цикла работы самой внутрикорковой системы возвратного торможения.
Таким образом, если в норме склонность соматической коры к ритмике проявляется в генерации веретена, то в других условиях (патология, фармакологические воздействия) вместо веретена может развиваться другой ритмический процесс, например, множественные повторения циклов торможения (генерация волн комплекса пик-волна может стать авторитмичной).
Уже отмечалось, что медленные волны, регистрируемые в коре на стимуляцию неспецифического таламуса, будучи сходны с petit mal — эпилептической активностью— по форме, частоте и топографии, в то же время отличаются от нее не только неспособностью к авторитмии, но и отсутствием синхронных с волнами судорог.
Вот почему теперь логично рассмотреть еще одну модель активности petit mal — эпилепсии, более близкую к истинной.
Нами была предложена такая модель petit mal — эпилепсии (Петухов, 1968; Гусельников, Петухов, 1971), которая подразумевала получение длительной спонтанной ритмической активности пик-волна при помощи применения специального подбора хлоралозно-барбитурового наркоза. Развитие такого смешанного наркоза протекает в несколько стадий. Вследствие того, что нембутал обладает меньшим латентным периодом действия по сравнению с хлоралозой, первая фаза наркоза, обусловленная лишь действием нембутала, ничем не отличается от обычного барбитурового наркоза и сопровождается развитием веретен как в коре, так и таламусе. Затем по мере проявления действия хлоралозы в различных структурах, и прежде всего в головке хвостатого ядра, появляется еще неустойчивая низкочастотная ритмика, В это время спонтанные сонные веретена, характерные для барбитурового наркоза, снижают частоту собственной ритмики и становятся более растянутыми во времени.
Когда выраженность волн в хвостатом ядре становится четкой, они начинают проявляться и в коре; причем прежде всего в зрительной, а затем и в других областях. Наступает 3-я стадия наркоза, когда практически вся кора захватывается синхронной ритмической активностью. В это время появляются спонтанные ритмичные подергивания сначала мимической мускулатуры, а потом и мышц конечностей. Они синхронны с ритмической активностью и со временем усиливаются, пока, наконец, не переходят в судороги, типичные для чистого хлоралозного наркоза. Все эти стадии занимают в общей сложности от 4 до 8 ч после введения животному смеси препаратов (50 мг/кг хлоралозы+10 мг/кг нембутала). Для нас наиболее интересна была фаза, на которой волны хорошо выражены в коре, но еще не сопровождаются судорожными движениями. Поэтому мы задерживали развитие наркоза на этой стадии введением небольшой дополнительной дозы барбитуратов (5 мг/кг нембутала).
Регистрируемая на этой стадии электрическая активность коры представляла собой низкочастотую (3—4 Гц) ритмику, состоящую из куполообразных поверхностноотрицательных волн, перемежающихся иногда небольшими пиками. Наибольшей амплитуды эти волны достигают в зрительной коре, где пик часто развивается не в промежутке между волнами, а на нисходящей ее части.
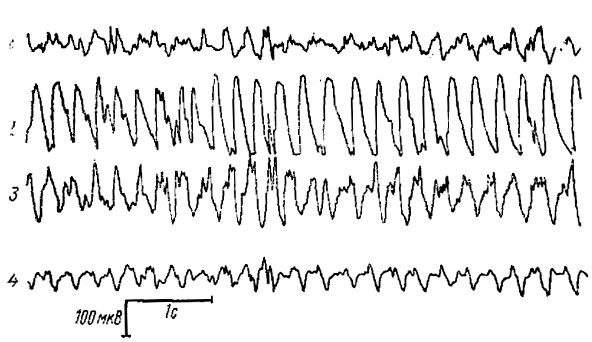
1 — отведение от centrum medianum, 2 — от латерального края задней сигмовидной извилины, 3 — от головки хвостатого ядра, 4 — от ассоциативной коры вдоль латеральной извилины
На стадии хорошо выраженных спонтанных волн вся кора «работала» как единое целое. Иногда при отведениях от зрительной и соматомоторной коры наблюдалась полная синфазность волн, вплоть до мелких деталей. Хотя в большинстве случаев корреляция активности была более слабой, достаточно четкая синфазность проявлялась на значительных корковых площадях. Между отведениями от коры (особенно соматомоторной) и от хвостатого ядра наблюдались противоположные отношения также прослеживающиеся вплоть до мелких деталей (рис. 5). Такая противофазность не может быть объяснена простым физическим «забросом» потенциала, поскольку волны в коре и в хвостатом ядре, как уже отмечалось, появляются не одновременно. Причины, обусловливающие устойчивые противофазные отношения, еще предстоит объяснить. Сейчас мы можем только сказать, что такая корреляция изменений потенциала между хвостатым ядром и соматомоторной корой наблюдается и во время вызванной активности типа пик-волна.
Чрезвычайная генерализация рассматриваемой спонтанной ритмики, значительная синфазность активности в удаленных друг от друга точках коры, которая проявляется даже в мелких деталях, заставляют предположить наличие единого ведущего центра этой ритмики, расположенного вне коры. Этот центр должен быть, во- первых, достаточно компактен и, во-вторых, обладать широкими проекциями ко всей коре. Исходя из данных о вызванной активности типа пик-волна, логично предположить, что этот центр расположен в неспецифическом таламусе. Однако наши данные показали, что самые разнообразные ядра как неспецифического, так и специфического таламуса проявляют на фоне спонтанных волн очень слабую корреляцию с корковой активностью; отводимая от них ЭЭГ часто не имеет ничего общего с корковыми отведениями. То же наблюдается и у человека (Rossi et al., 1968). Эти факты ставят под сомнение возможность нахождения водителя спонтанной ритмики пик-волна среди таламических ядер. Единственной из рассмотренных структур, в которой такая корреляция активности всегда была четкой и волны возникали раньше, является хвостатое ядро. Наши данные, позволившие приписать ему роль водителя, были основаны сначала только на одном сравнении электрической активности различных мозговых структур во время хорошо выраженных спонтанных волн.
Наличие прямых каудато-кортикальных путей долгое время отрицалось, хотя электрофизиологические данные свидетельствовали о наличии таких связей (Бутхузи, 1970; Buchwald, Hull, 1967; Хасабов, Кураев, 1968, и многие другие).
В морфологической же литературе имеются данные о непрямых, переключающихся каудато-кортикальных проводящих системах (Загер, 1962; Szabo, 1970 и др.). Допускаются только опосредованные связи с корой через бледный шар и ядра неспецифического таламуса. Как показали наши данные, передача ритмической активности от хвостатого ядра через неспецифический таламус маловероятна, так как ядра последнего проявляют чрезвычайно слабую корреляцию электрической активности пик-волна как с корой, так и с хвостатым ядром.
Однако в последние годы были получены новые данные о прямых связях хвостатого ядра с корой больших полушарий. Было достоверно выяснено, что хвостатое ядро имеет прямые связи с двигательной, орбитальной, лимбической и обеими сомато-сенсорными зонами коры больших полушарий (Ермоленко, 1969; 1973). Это уже дает достаточные основания полагать, что хвостатое ядро может быть водителем ритма пик-волна в коре больших полушарий. Интересно отметить и тот важный факт, что волокна, поступающие в кору из хвостатого ядра, группируются больше всего вокруг мелких нейронов трех нижних слоев коры, нейронов, являющихся наиболее возможными «кандидатами» на роль тормозных вставочных нейронов, осуществляющих в коре возвратное торможение (Ермоленко, 1973).
Возможен, видимо, и еще один механизм широкой генерализации активности пик-волны в коре. Хорошо известны прямые связи хвостатого ядра с лобной корой в районе орбитальной и надглазничной извилин (Harman et al., 1954). Однако эти же участки коры, видимо, посылают широкие проекции ко всем остальным отделам новой коры.
Основываясь на этом, мы в свое время построили гипотетическую схему отношении между мозговыми структурами во время генерации спонтанной активности типа пик-волна (Гусельников, Петухов, 1970). Хвостатое ядро по этой схеме служит первичным очагом ритмики, которая через орбитальную кору распространяется на всю остальную кору. Такая структура является одновременно и компактной и быстропроводящей.
Для проверки этого предположения были проведены опыты с разрушением орбитальной коры на фоне спонтанных волн. Двустороннее разрушение орбитальной коры сопровождалось исчезновением спонтанных волн в остальной коре при сохранении их в хвостатом ядре. Морфоконтроль показал, что разрушалась именно орбитальная кора. Таким образом, данные подтвердили нашу гипотезу о ведущей роли системы: хвостатое ядро – орбитальная кора в генерации спонтанных волн по всей коре, где хвостатое ядро является водителем этого ритма.
Поллен (Pollen, 1968) полагает, что в генерализации активности пик-волна по коре больших полушарий участвует мозолистое тело, а также, вероятно, и фронто-орбитальная область коры, которая получает прямые проекции из интраламинарных ядер таламуса и оказывает прямое или косвенное (например, через полосатое тело) влияние на двигательные и сомато-сенсорные области коры. Видимо, чтобы окончательно решить вопрос о широком вовлечении зон коры в активность, необходимы еще дополнительные исследования.
1, 3— отведение от соматосенсорной коры. 2 — отведение миограммы от одной из малых мимических мыши, 4 — от сгибателя передней контрлатеральной по отношению к отведению ЭЭГ конечности
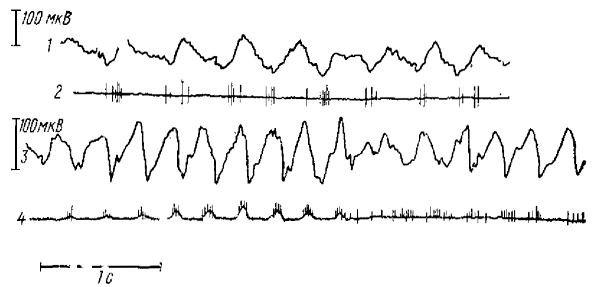
Рассмотрим некоторые другие черты спонтанной ритмики. На определенной стадии смешанного наркоза появляются ритмические, синхронные с электрической активностью слабые сокращения (вздрагивания) мышц животного. Как показали синхронные записи миограммы и ЭЭГ соматомоторной коры (рис. 6), вспышки мышечной активности приурочены к положительным фазам волн. Более четким волнам соответствует усиление мышечных сокращений (что выражается в искривлении линии миограммы). Эти ритмические сокращения мышц чрезвычайно сближают наблюдаемую ритмику с клинической картиной petit mal — эпилепсии. Кривые показывают, что амплитуды волн и двигательной реакции находятся в прямой зависимости. Более мощным волнам соответствует уже не только пачка мышечных спайков, но и сдвиг изопотенциальной линии, вызванный артефактом движения. Уже отмечалось, что эти движения очень чувствительны к дополнительным дозам барбитурата. Так, добавление всего 3—5 мг/кг нембутала, которое очень слабо изменило бы поведение, животного в норме, на фоне смешанного наркоза вызывает резкое подавление этих движений. Отсюда ясно, что в опытах Поллена вызванные волны никогда не сопровождались какими-либо двигательными коррелятами, так как он применял нембутал в дозировке 15—20 мг/кг. Добавление животному 10—15 мг/кг нембутала в наших опытах вызывало не только прекращение ритмических вздрагиваний, но и исчезновение самих волн, возвращая смешанный наркоз к его начальной стадии. Подобное воздействие барбитурата позволяет предположить, что любая искусственная активация животного будет давать противоположный эффект, а именно делать ритмику более четкой и усиливать ее миоклонические корреляты. Действительно, как болевое раздражение, так и высокочастотная стимуляция стволовой ретикулярной формации вызывают резкое увеличение волн и усиление связанных с ними судорожных явлений.
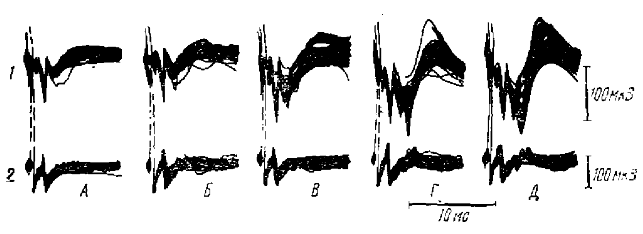
Гусельникову и Петухову, 1971)
1 — отведение от проекционной зоны зрительной коры, 2 — от латеральное коленчатого тела
В заключение сравним спонтанную и вызванные ритмики типа пик-волна. Между ними, без сомнения, существуют некоторые различия. Прежде всего, спонтанная ритмика в отличие от вызванной характеризуются способностью к авторитмии. Вероятно, это связано с тем, что в случае спонтанных волн активируется вся система, обеспечивающая ритмику, в том числе и ее водитель, в то время как при вызванной ритмике активируется, очевидно, лишь последнее звено этой системы — механизм внутрикоркового возвратного торможения. Рассматриваемые виды ритмики различаются также по корковой типографии. Если вызванные волны приурочены лишь к ростральным отделам и никогда не встречаются в затылочной области коры, то спонтанные волны регистрируются по всей коре и лучше всего выражены в зрительной области. Однако между этими двумя видами электрической активности гораздо больше сходства, чем различий. Сходны форма н частота волн во время установившейся ритмики. Частота спонтанных волн довольно строго ограничена пределами 3—3,5 Гц и лишь на поздних стадиях смешанного наркоза повышается до 4—4,5 Гц. При воздействии спонтанных волн с первичным ответом в зрительной коре наблюдается подавление первичного ответа в начале волны (рис. 7). Это говорит в пользу тормозной природы спонтанных волн и подтверждается корреляцией максимумов нейронной активности в коре с положительными фазами спонтанной ритмики, которые разделены промежутками полного молчания, занимающими большую часть длительности волны. Полученные данные свидетельствуют о внутрикорковом характере этого торможения (рис. 7). Так, вызванный потенциал в латеральном коленчатом теле (нижняя запись в каждой осциллограмме) практически не испытывает подавления в ходе волны, в то время как вызванный корковый потенциал изменяется значительно.
Приведенные факты, видимо, могут свидетельствовать о том, что как в случае вызванных, так и в случае спонтанных волн мы имеем дело с процессами одинаковой природы — с проявлением активности внутрикорковой системы возвратного торможения.
О внутрикорковой природе этого торможения косвенно могут свидетельствовать данные о наличии сходной судорожной активности, наблюдаемой на препарате изолированной от подкорковых связей коры (Хананашвили, Буракова, 1969). Кроме того, система возвратного торможения, как показали данные нашей лаборатории, является, видимо, элементарным механизмом торможения нейрона в любых нервных структурах животных разного уровня эволюционного развития. Видимо, в силу этого можно полагать, что нейрональные механизмы активности пик-волна могут быть сходны начиная с переднего мозга рыб и кончая новой корой млекопитающих. Разница, возможно, заключается лишь в системе провокации этих нейрональных механизмов.
Существует представление, что включение системы возвратного торможения, отражающейся в виде медленной волны в ЭЭГ, способствует стабилизации малого припадка и препятствует его трансформации в большой эпилептический приступ (Dalby, 1969).