В настоящее время термин «вторичный ответ» (ВО) приобрел некоторую неопределенность. В свое время поводом для разделения первичных (или специфических) ответов от вторичных (неспецифических) ответов послужили четкие экспериментальные данные. ПО коры связывали со специфической афферентацией коры, а ВО — с неспецифической афферентацией из ретикулярной формации.
В 1936 г. Дербишайр и сотрудники (Derbishire et al.) описали у наркотизированной кошки генерализованную по коре двухфазную негативно-положительную реакцию с большим латентным периодом от 40 до 80 мс, возникающую на раздражение седалищного нерва. Такая же реакция была описана еще ранее Правдич-Неминским (1913).
В 1939 г. Форбс и Морисон (Forbes, Morison), изучив свойства этой реакции, возникающей по времени позднее ПО, предложили для нее термин «вторичный разряд». Эта реакция известна в литературе и как «ответ Форбса».
Два года спустя Морисон и сотрудники показали, что такой же «вторичный разряд» возникает в коре полушарий при раздражении различных уровней мозгового ствола. В последующих работах многих исследователей было выяснено, что генерализованные корковые реакции — вторичный ответ — возникают на раздражение любого участка ретикулярной формации. Основными отличиями ВО от ПО считались длинный латентный период ВО, его слабая устойчивость к повторным раздражениям, широкое распространение по коре. Проявление вторичного ответа при глубоком нембуталовом наркозе, внешнее отличие ВО от позитивно-негативного комплекса волн ПО.
Позже указанная реакция была выделена из других видов вторичных реакций (Кулланда, 1964) в самостоятельную вторичную генерализованную реакцию ретикуло-стволового происхождения.
В последующие годы было найдено большое количество реакций, имеющих существенные отличия как от классического ПО, так и от вторичного ответа — ответа Форбса. В литературе описано много различного рода ВО под разными названиями: ранних, ассоциативных, поздних, отставленных, неспецифических, непервичных и т. д. Все они причислялись к ВО и связывались с ретикулярной системой. С другой стороны, критерии определения вторичного ответа, о которых мы говорили выше, были уже неприменимы ко многим ВП. Так, например, были открыты нового вида генерализованные неспецифические реакции, или локальные реакции не ясного еще генеза, с латентным периодом более коротким, чем у ПО, а также равным ему или чуть его превышающим. Были зарегистрированы ВО, по форме сходные с ПО, и специфические ответы, по форме не отличающиеся от ВО. То же касается критерия распространения реакций по коре. Были найдены ВО, не регистрирующиеся за пределами проекционной зоны, или ограниченные другими определенными зонами коры, в то время как ПО, видимо, регистрируются шире по коре, чем считалось ранее. Были описаны ВО и у бодрствующих животных, так что и критерий применения наркоза для регистрации ВО оказался необязательным. Было также показано, что ряд вторичного ответа связан со специфической афферентацией.
Эти и многие другие факты заставили ряд исследователей поставить вопрос о пересмотре классификации вызванных ответов коры и принципов их разделения на ПО и ВО (Кратин, 1971; Батуев, 1971, и др.).
Однако, как нам кажется, еще рано пытаться ввести какую-либо новую классификацию ВП коры. Это связано прежде всего с тем, что у нас еще нет точных критериев для такой новой классификации. Анализ фактического материала настолько еще противоречив, что он не может быть положен в основу какой-либо приемлемой классификации, тем более что генез большого количества описанных ВП коры вообще еще не ясен. Поэтому термины ПО и вторичного ответа рационально сохранить, понимая под ПО тот характерный классический комплекс ВП проекционных зон анализаторов. Термин ВО полезно, видимо, также пока сохранить для всех видов других потенциалов, каковы бы ни были их латентный период, зона регистрации и другие свойства. Исторически, с нашей точки зрения, это более приемлемо, пока не будут найдены четкие критерии классификации ВП.
Естественно, что термин «вторичный» надо понимать крайне условно, акцентируя основное внимание при описании ВО на их генезе и функциональных свойствах. В связи с этим мы считали целесообразным в общих чертах сохранить для ВО классификацию К. М. Кулланды (1964, 1968), позволяющей провести некоторую грань между ВО разного происхождения.
Вторичные локальные ответы
Ранние отрицательные реакции (POP). POP обнаружены К. М. Кулландой и В. А. Черниговским (1957, 1959, 1968) у кошек под легким хлоралозным наркозом. POP возникают в ответ на стимуляцию сенсорных нервов, вслед за ПО.
Таблица 1. Полярности первой фазы ПО, POP, POP-I, POP-II при раздражении седалищного нерва и ядер таламуса (по Кулланде, 1969)
| Зоны отведения потенциалов в коре, определяемы при стимуляции седалищного нерва | Стимулируемая структура | ||||
|---|---|---|---|---|---|
| n. ischiadicus | n. VLP thalami | n. VA thalami | |||
| Первичный ответ | + | + | ± | ||
| Постсинаптический POP | – | – | + | ||
| Пресинаптический POP-I | – | – | + | ||
| Пресинаптический POP-II | – | + | – | ||
| Полярность I фазы потенциалов | |||||
Как и ПО, POP регистрируется от локальных участков по соседству с зоной ПО, но зона POP не окружает зону ПО. Фокусы максимальной активности ПО и POP расположены на расстоянии нескольких миллиметров друг от друга, а между ними расположена переходная зона, где регистрируются ответы переходной формы. POP представляют собой в основном отрицательные колебания с латентным периодом на 2—3 мс больше, чем латентный период ПО.
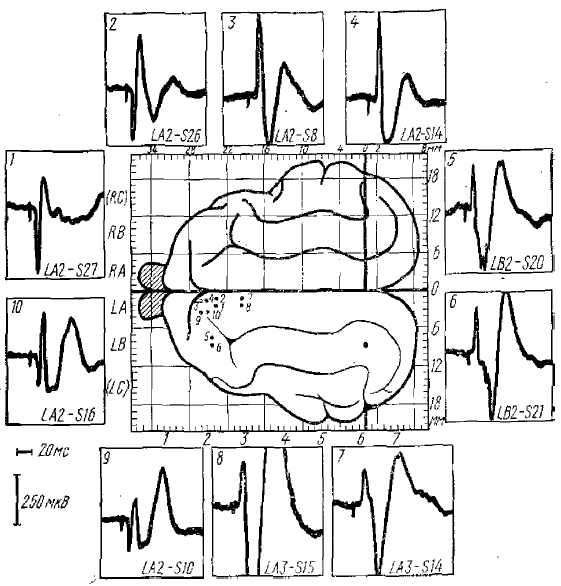
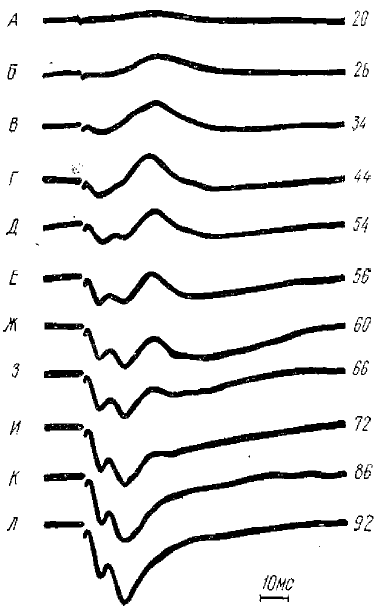
Первичные ответы 1—2 — коротколатентные («классические»); 9—10 — длиннолатектные. Ранние отрицательные реакции: 3—-4 — постсинаптические; 5—6 — пресинаптические I типа; 7—й — пресинаптические II типа. Отклонение луча вниз соответствует положительному колебанию потенциала. В центре — схема мозга кошки с системой координат стереотаксического атласа. 1—10 — на схеме точки отведения потенциалов. Обозначения внизу каждого кадра — стереотаксические координаты отведения
POP найдены лишь при стимуляции соматической и слуховой систем (рис. 1).
В результате последующих исследований К. М. Кулланда пришел к выводу, что можно зарегистрировать три различных вида POP, изучение генеза которых позволило подразделять их на постсинаптические POP и пресинаптические POP I и II типов. Все эти виды POP возникают в I и II проекционных зонах сомато-висцеральной чувствительности и слуховых проекционных зонах.
POP различаются, во-первых, латентными периодами (пресинаптические POP имеют одинаковые латентные периоды, а постсинаптические — больший), а также амплитудой начальной отрицательной фазы; последняя у постсинаптических POP больше, чем у пресинаптических.
Основное отличие, как отмечает К, М, Кулланда (1969), заключается в том, что на пре- и постсинаптические POP по-разному влияют синаптоактивные химические вещества (стрихнин, ГАМК, метразол и др.) (рис. 2). Их применение вызывало изменения у одних POP и не влияло на другие. Это и легло в основу их разделения на пост- и пресинаптические.
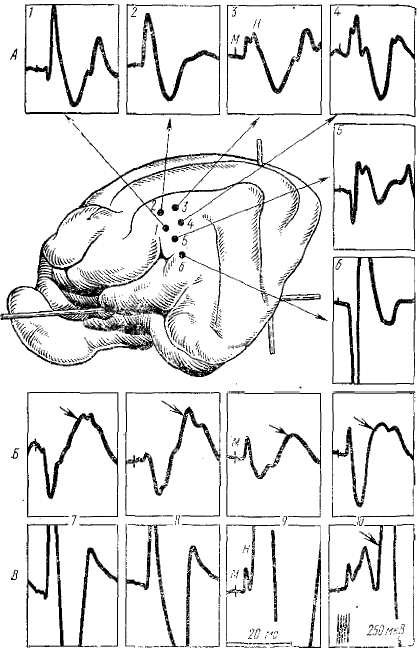
1, 2, 7, 8—постсинаптические POP; 3, 4, 9, 10 — пресинаптические POP; 5—6 - первичные ответы. Артефакт cooтветствует моменту звукового стимула
В основу разделения пресинаптических POP на два вида легли два рода фактов: различие мест их регистрации и полярности на раздражение определенных таламических ядер. Так, пресинаптический POP I типа начально отрицателен при раздражении VLP (nucl. ventralis postcrolateralis) ядра таламуса и начально положителен при раздражении VA (nucl. centralis anterior) ядра таламуса. Для пресинаптического POP II типа начальные полярности при раздражении тех же ядер прямо противоположны (табл. 1).
К. М. Кулланда полагает, что начальная отрицательная фаза пресинаптических POP связана с возбуждением пресинаптических структур (с входящими в поверхностные слои волокнами), в то время как отрицательная фаза постсинаптических POP является ВПСП апикальных дендритов пирамидных нейронов коры.
POP не являются чистым «лабораторным феноменом», связанным с наркозом или какими-либо условиями эксперимента, так как они регистрируются и в хронических условиях опыта с вживленными электродами, что, по мнению К. М. Кулланды, говорит о том, что «РОР являются отражением неких общих принципов функциональной организации корковых проекций анализаторных систем».
Однако сказать что-либо об информационном значении POP пока не представляется возможным. К. М. Кулланда (1964) полагает, что они могут отражать связь специфических корковых проекций с соответствующими ассоциативными зонами коры, а также связь коры с ядрами таламуса, мезенцефалической частью РФ и с мозжечком. Доказательств этому положению пока нет.
Вызванные потенциалы в коре с начальной отрицательностью были зарегистрированы и другими исследователями (Нарикашвили, 1956; Steriade, 1964; Гмыря-Нови, 1968, Гмыря-Нови, Васечко, 1970 и др.). Насколько POP и отрицательные ответы коры, найденные этими авторами, сходны, сказать трудно.
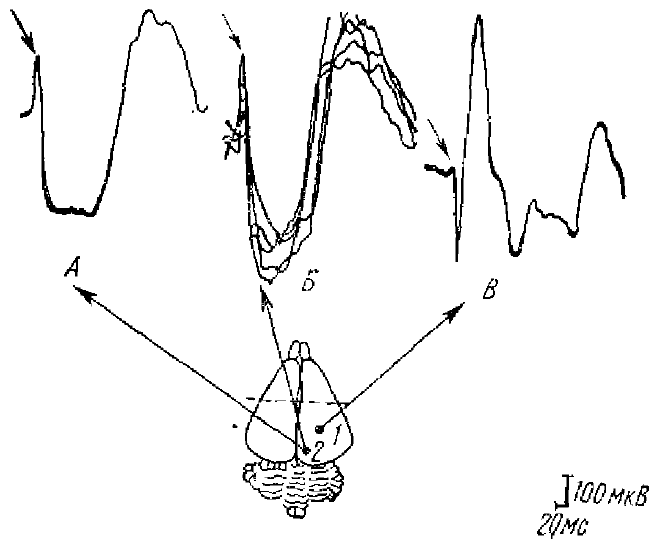
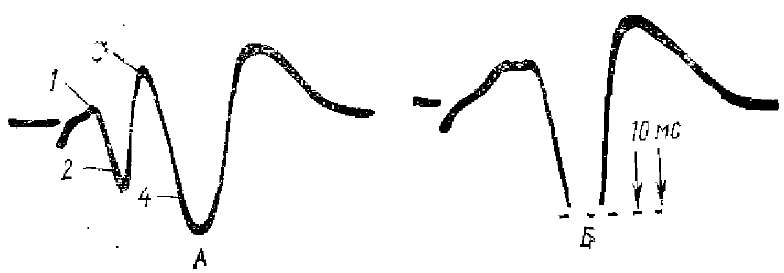
На А, Б и В — стрелкой отмечен ЭОК: 1 — ФМА вызванного ответа в зрительной коре, 2 — фокус ЭОК
Экстрапервичный отрицательный компонент зрительного потенциала (ЭОК) был описан в 1971 г, Ф. А. Ата-Мурадовой и Л. М. Чуппиной. Он регистрируется перед ПО зрительной зоны коры, Нисходящая фаза этого ответа в ФМА ПО сразу переходит в позитивный компонент ПО (рис. 3). Этот отрицательный компонент ПО регистрируется не только в зрительной коре, но и за ее пределами. Латентный период ЭОК 20 мс, на некотором расстоянии от ФМА амплитуда и длительность ЭОК растет. ЭОК более устойчив к сильным дозам наркоза, когда ПО резко подавлен. Таким образом, авторы подчеркивают, что ЭОК отражает приход в кору самого быстрого потока из всех восходящих возбуждений от сетчатки, это возбуждение более генерализованное, чем ПО, и оно самое устойчивое к большим дозам наркоза. Приведенные авторами исследования показали, что ЭОК является постсинаптическим потенциалом.
Вопрос о путях проведения в кору зрительной информации, вызывающей ЭОК, остается пока открытым. Авторы полагают, что он связан с проведением либо по прямым путям, связывающим сетчатку с корой, либо это должны быть пути, по крайней мере, малосинаптические. Хотя прямые пути из сетчатки в кору известны (Дзугаева, 1958, 1960; Бирючков, 1963), но поскольку они состоят из тонких волокон, вряд ли могут определять появление ЭОК в коре в связи с малой скоростью проведения по ним импульсов.
Длиннолатентные «первичные» ответы (ДПО). Этот термин введен К. М. Кулландой (1968) для ответов, крайне напоминающих ПО, но, по его мнению, между ДПО и ПО есть и существенные различия (см. рис. 1). Это касается прежде всего различий в их латентных периодах в 2,4 мс при раздражении седалищного нерва генерирующего ДПО, что свидетельствует о дополнительных синаптических переключениях потока импульсов в таламокорковой системе.
ДПО генерируются не в фокусе максимальной активности для ПО, но имеют ограниченную зону отведения. Кулландой найдена существенная разница в инверсии фаз ДПО по сравнению с ПО при погружении электрода в кору. Особенностями этого ответа является отсутствие его инверсии в глубоких слоях коры и ряд своеобразных изменений положительной и отрицательной фаз.
Таким образом, описанные три группы ответов с большой вероятностью могли бы быть отнесены к ПО коры по латентным периодам, по характеру специфической информации, их вызывающей, и по ряду других своих свойств. Однако все они имеют и существенные отличия от классических ПО. Такой широкий спектр различных по генезу ВП в проекционных зонах анализаторов еще раз подчеркивает сложность функциональной организации последних и гетерогенность приходящих к ним возбуждений, что неоднократно отмечалось П. К. Анохиным и его сотрудниками.
Вторичные поздние отрицательные реакции, найденные К. М. Кулландой в 1964 г., возникают при хлоралозном наркозе в I сенсомоторной зоне с длинным латентным периодом порядка 50—100 мс в виде медленной отрицательной волны амплитудой в 150—200 мкВ. Близкие по форме и свойствам реакции были зарегистрированы и другими исследователями в слуховой коре кошки на световые раздражения. К. М. Кулланда полагает, что поздние отрицательные реакции имеют ассоциативный характер и отражают поздние этапы переработки информация.
Вторичные ассоциативные ответы (ВАО). Ассоциативные области коры больших полушарий, в отличие от первичных проекционных зон, характеризуются тем, что в них возникают ВП, имеющие сходные латентные периоды, форму и продолжительность при самых различных периферических раздражениях: световых, звуковых, соматосенсорных. Эти ВП получили название вторичных ассоциативных ответов — ВАО. В литературе можно встретить для этих ответов и другие термины: «вторичные», «ассоциативные», «конвергентные» и др.
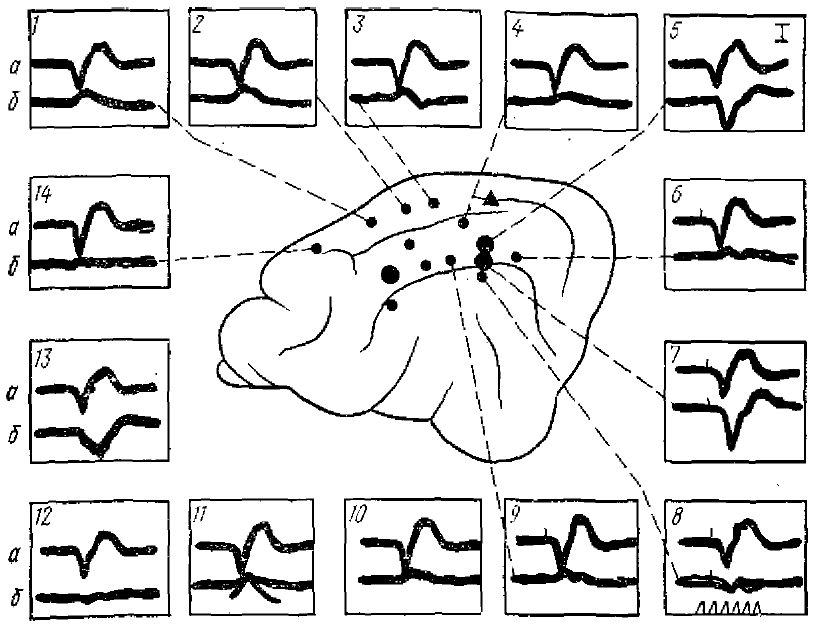
1—14 — одновременная регистрация ПО (а) и ВАО (б). На схеме мозга в центре; черные точки — пункты регистрации ВАО; треугольник — ПО. Время — (кадр 8) 50 Гц; масштаб усиления (кадр 5) — 150 УКБ
ВАО имеют значительно больший латентный период, чем ПО, они не регистрируются в проекционных зонах анализаторов. Часто их форма сходна с формой ПО, однако они отличаются большей продолжительностью и величиной положительной фазы (рис. 4). Описаны ВАО, имеющие и другую форму колебания, состоящие из одного позитивного или одного негативного колебания или негативно-позитивного комплекса (Amassian, 1953; Дуринян, 1965, 1966; Дуринян, Полякова, 1967, и др ).
ВАО в пределах раздражения одной модальности возникают на сигналы разного характера. Так, например, в соматической области коры ВАО возникают при раздражении любого участка тела как ипси-, так и контралатеральной сторон.
Некоторые исследователи полагали, что вторичный ответ Форбса и ассоциативные ответы — явления одного порядка. Оба они имеют большой латентный период, большую «утомляемость» при повторных раздражениях и сходные области коры, где они лучше всего выражены. Однако между ними есть существенная разница: вторичный ответ Форбса регистрируется в условиях глубокого нембуталового наркоза, в то время как ВАО возникают при хлоралозном наркозе или у ненаркотизированных животных.
Дальнейшие исследования ряда авторов показали, что ответы ассоциативных областей коры должны быть выделены в особую группу ВП, так как они имеют ряд принципиальных и свойственных только им особенностей. Это касается прежде всего особенностей локализации ВАС) в пределах ассоциативных зон коры, их коркового электрогенеза и подкорковых механизмов, лежащих в основе их генерации.
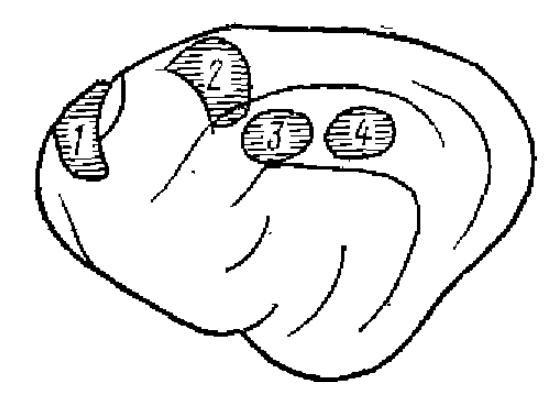
1 — вокруг крестовидной борозды, 2— передней латеральной извилины, 3 — переднего участка средней суирасильвиевой извилины, 4 — заднего участка средней супрасильвиевой извилины
На основании регистрации ВАО, возникающих на разные периферические раздражения в ассоциативной коре животных, выделено несколько зон (отделяющихся от первичных проекционных зон участками коры, слабо или вовсе не отвечающими на раздражения) (рис. 5).
Так, например, у кошки выделено несколько ассоциативных зон. К ним относят переднюю часть краевой извилины, передний и задний участки средней части супрасильвиевой извилины, моторную кору с гиганто-пирамидным полем, прореальную извилину и орбитальную кору. В этих зонах отмечается конвергенция импульсации различной модальности: зрительной, слуховой, соматической. Одним из важных фокусов всех этих трех афферентаций является средняя супрасильвиевая извилина. Наиболее сложно построена фронтальная и околокрестовидная зона, где регистрируются хорошо выраженные ВАО практически на все виды стимуляции. По данным большинства авторов, в этих участках коры на все периферические раздражения возникают ВАО одинакового характера. Полагают (Thompson et al., 1963; Albe-Fessard et al., 1963), что при раздражении разных афферентных систем возбуждается одна и та же центральная подкорковая система, которая совершенно одинаково влияет на разные участки ассоциативной коры. Если это так, то тогда в кору поступают уже интегрированные на подкорковом уровне импульсы, которые для коры теряют свою специфичность — сигнальное значение.
Этот вопрос подвергся специальному анализу как методом изучения динамики ВАО, так и на нейронном уровне (Нарикашвили и сотрудники, 1967, Dubner et al., 1964, 1965; Айрапетьянц, Батуев, 1969; Батуев. 1970, и др.).
Была показана существенная разница в ВАО разных участков ассоциативной коры, которая выражалась в различной амплитуде и латентных периодах на разные модальности раздражителей. Так, были получены данные преимущественной проекции слуховой и зрительной систем в разных участках ассоциативных зон. Изучение взаимодействия раздражений различных модальностей выявило сложную динамику облегчения и блокады (окклюзии) между разными афферентными сигналами на уровне нейронов ассоциативных зон коры. Конечный результат зависел от того, какое раздражение было кондиционирующим и какое тестирующим. Полученные данные дали основание полагать, что механизм интеграции разных афферентных импульсов происходит на уровне коры, что свидетельствует о том, что к ассоциативной коре поступают не интегрированные на уровне подкорки импульсы, потерявшие, по мнению Альб-Фессар и других (Albe-Fessard, Fessard, 1963), свою специфичность, а импульсы, несущие определенную информацию о внешнем сигнале (Нарикашвили, 1967).
Позже, в 1971 г., Альб-Фессар, признавая факт истинной конвергенции импульсов от различных афферентных систем на нейронах ассоциативных зон коры, полагает, что такие мультисенсорные области коры (как и подкорковых структур) необходимо разделить на две группы: неспецифические и специфические конвергентные области. Она считает, что потоки импульсов, приходящие в неспецифические конвергентные области, не содержат в себе локального сигнала, что не позволяет надежно анализировать длительность стимула. Реакция нейрона в этом случае зависит только от интенсивности стимула.
Однако существуют, по ее мнению, области коры (и подкорки), где конвергируют гетеромодальные афференты специфического типа и где, следовательно, могут происходить более топкие интермодальные ассоциации. Это — ассоциативные области коры, куда она относит супрасильвиевую извилину, первичную зрительную кору (так как сюда приходят и специфические слуховые афференты), соматическую область II, орбитальную кору, моторную кору. Таким образом, по Альб-Фессар, одни ассоциативные области коры получают только сенсорную неспецифическую афферентацию, а другие — мультисенсорную специфическую афферентацию.
Эти данные, полученные на нейронном уровне, значительно усложнили проблему определения ассоциативных зон и функционального значения мультисенсорной конвергенции (Айрапетьянц. Батуев, 1969; Полянский и др., 1970; Дуринян. 1970; Альб-Фессар, 1971; Воронин и др., 1971).
До настоящего времени не существует еще окончательного мнения относительно тех путей, по которым афферентные сигналы поступают в ассоциативную кору. Исходя из величин латентного периода ВАО импульсы приходят в ассоциативные зоны коры, пройдя большое количество синаптических переключений. Вначале полагали, что импульсация, вызывающая ВАО, распространяется транскортикально из первичных проекционных зон коры и что ассоциативные области лишены периферической проекции. Однако опыты с изоляцией проекционных зон коры (путем отделения их от остальных участков коры) и опыты с их экстирпацией показали, что ВАО в ассоциативных областях коры продолжали регистрироваться.
Было высказано предположение, что пути, идущие к первичной сенсорной зоне и к ассоциативной коре, разделены чуть ли не с рецептора (Albe-Fesard, Fesard, 1963). Однако экспериментально было выяснено, что удаление части ретикулярной формации всего промежуточного мозга (в том числе и срединного центра) не препятствует появлению ВАО, если остается интактной часть промежуточного мозга, переключающая импульсы к первичным зонам коры (Bignal et al., 1966; Dubner, Brown, 1968, и др.). Например, если при разрушении всего промежуточного мозга остается интактным наружное коленчатое тело, ВАО на свет также остаются, но полностью исчезают на другие модальности раздражений. Таким образом, импульсы, идущие к ассоциативной и первичной проекционной коре, идут вместе до релейного ядра промежуточного мозга. Однако где они расходятся, еще не ясно.
Хотя и известно много путей от сетчатки к разным подкорковым образованиям, но, как отмечает С. П. Нарикашвили (1970), принимая во внимание вышеуказанные данные, эти пути должны проходить через наружное коленчатое тело. В остроумных опытах со слабым электрическим раздражением разных слоев наружного коленчатого тела С. П. Нарикашвили с сотрудниками, было найдено, что верхние два слоя коленчатого тела посылают импульсы к первичной проекционной зоне зрительной коры, а нижний слой — к ассоциативной, через латеральное ядро таламуса. При повреждении нижнего слоя исчезал ВАО, а при повреждении верхних слоев — ПО. Таким образом, одна и та же зрительная информация по более коротким путям достигает проекционной зоны коры, а по более длинному пути — ассоциативной области. Об участии наружного коленчатого тела в проведении ассоциативного ответа на свет в двигательную область коры говорят данные, полученные Л. В. Черенковой (1973). О роли релейных ядер в происхождении ВАО указывают и другие авторы (Imbert, 1966; Bignail, 1967, и др.). А. С. Батуев, анализируя пути проведения световой и звуковой информации в сенсомоторную кору кошки, вызывающих ВАО (область гиганто-пирамидных полей) пришел к заключению, что ответственной за проведение является самостоятельная таламо-кортикальная система, отличная по своим свойствам от специфических афферентных систем. Проведение в этой системе блокируется нембуталом и облегчается хлоралозой. Он полагает, что основную роль в проведении ВАО играет таламическая ассоциативная система (дорсомедиальные и частично заднелатеральные ядра), получающая волокна от внутреннего и наружного коленчатых тел.
Надо отметить, что еще в 1939 г. Бюзе и сотр. (Buse et al.) также пришли к выводу, что ВАО связаны с возбуждением, приходящим в кору от ассоциативных ядер таламуса. По мнению авторов, возбуждение на уровне таламуса передается от специфических ядер к ассоциативным, а нейроны последних передают его в определенные зоны коры.
Высказывается также предположение, что может иметь место и непосредственная активация некоторых ассоциативных ядер таламуса импульсацией, поступающей прямо от рецепторов по тонким волокнам (Bignail, 1967). О прямой активации ассоциативных ядер говорят данные Нарикашвили и др. (1971) и Агилара и др. (Aguilar et al,. 1962). Однако надо отметить, что пока группа ассоциативных ядер таламуса слабо изучена и роль их еще далеко не ясна. В последние годы получены многочисленные данные о сложности связей этих ядер с корой, с релейными и неспецифическими ядрами таламуса. Огромная и противоречивая литература этого вопроса требует критического подхода в ее обобщении и значительной экспериментальной проверки. Особая трудность в обобщении материала связана также с тем, что исследователи пользуются различной терминологией и методическими приемами в изучении одних и тех же ВАО.

1 — первичный ответ ПО в фокусе максимальной активности, 2—на периферии зоны представительства седалищного нерва в проекционной коре, 3 —зона молчания между проекционной и ассоциативной корой, 4—8 — РАО различных точек ассоциативной коры, 9 — отсутствие характерного РАО на периферии ассоциативной коры, 10 — сравнение латентных периодов ПО и РАО, равных 10 мс. На схеме мозга цифры соответствуют точкам отведения ПО и РАО
Кроме длиннолатентных ВАО рядом авторов были описаны и коротколатентные, или ранние ассоциативные, ответы (РАО) (Дуринян, Полякова, 1965; Полякова, 1965, 1967, 1970, 1972, и др.) (рис. 6). РАО возникают в разных пунктах теменной коры. Коротколатентный РАО представляет собой отрицательное или положительно-отрицательное колебание; негативная волна преобладает над позитивной, его латентный период равен таковому ПО. РАО возникает независимо от длиннолатентного ВАО и регистрируется как у неанестезированного, так и у анестезированного хлоралозой животного. Коротколатентное колебание было обозначено как ранний комнонент ассоциативного ответа (РКАО), а длиннолатентный ответ—как поздний компонент ассоциативного ответа (ИКАО) (рис. 6).
Полагают, что РКАО обусловлен передачей активности из проекционных отделов анализаторов по интеркортикальным волокнам (Баклаваджян, 1965; Нарикашвили, 1969). Однако Р. А. Дуринян и А. Г. Полякова (1965); А. Г. Полякова (1970) показали возможность автономного возникновения РКАО вне зависимости от проекционных областей. Опыты А. Г. Поляковой показали, что РКАО связан с возбуждением нервных элементов ассоциативной коры и формируется залпом импульсов, идущим по толстым волокнам из специфических ядер таламуса. Исходя из тех факторов, что для РКАО в ассоциативной коре отсутствует соматотопия, ФМА и инверсия при проходке электродом коры, Дуринян и Полякова полагают, что его генез связан с особенностями строения ассоциативной коры — наличием в последней терминалей специфических волокон во всех слоях коры и особенно их аксодендритных связей. С последним они связывают преобладание у РКАО отрицательной фазы.
Все сказанное выше показывает, что ВАО в разных зонах ассоциативной коры обладают своими особенностями. В связи с этим, надо полагать, их генез имеет определенные различия. Эти различия связаны прежде всего со спецификой афферентации данной ассоциативной зоны из подкорковых структур, с особенностями синаптических окончаний этих афферентов на нейронах данной области, а также с особенностями внутринейрональных связей данного участка ассоциативной коры. Поэтому в исследованиях последних лет наметилась определенная тенденция более тщательного анализа ВАО конкретных ассоциативных областей коры. Такой анализ связан прежде всего с изучением синаптических процессов, лежащих в основе генерации ВАО (Казаков, 1970; Батуев, 1970, и др.).
Влияние первичных, проекционных областей коры на ассоциативную кору
Если сейчас не вызывает сомнения тот факт, что ВАО возникают в результате афферентной импульсации из подкорковых центров (а не за счет импульсации от проекционных зон коры), то вопрос о влиянии проекционных полей, на ассоциативную кору является предметом изучения. Ряд авторов экспериментально показал, что ассоциативная кора находится под постоянным контролирующим влиянием первичных воспринимающих областей (Баклаваджян, 1965; Нарикашвили. 1967, 1968, 1969; Amassian, 1954; Buser, Borenstein, 1957; Казаков, 1968, и др.) и что эта связь односторонняя — от проекционных зон к ассоциативной коре.
Вторичные каллозальные ответы (ВКО). Известна важная роль ассоциативных волокон, составляющих значительную часть мозолистого тела, в парной деятельности больших полушарий головного мозга.
Еще в 1925 г. В. Я. Данилевским было показано, что локальное раздражение электрическим током пункта коры одного полушария вызывает ответ в симметричном пункте коры противоположного полушария. В последующих исследованиях ряд авторов подтвердили эти наблюдения и провели первые исследования каллозальных потенциалов (Curtis, 1940). Более детальный анализ был проведен Чангом (Chang, 1953) и затем Ройтбаком (1955).
Чанг показал, что ВКО возникает с коротким латентным периодом (0,5—1 мс) и по своей форме он сходен с ПО, причем сам ВКО возникает на участке коры противоположного полушария, не превышающем 4 мм². ВКО представляет собой двухфазное положительно-отрицательное колебание (рис. 7)
К несколько иным результатам пришел А, И. Ройтбак, который считает, что ВКО могут наблюдаться (при легком наркозе) не только в симметричном пункте противоположного полушария, но значительно шире — в несимметричных пунктах и даже в несоответствующей извилине коры. Таким образом, по А. И. Ройтбаку, транскаллозальные связи полушарий значительно более сложны и обширны и не соответствуют принципу связи «точка в точку». Расширение при определенном функциональном состоянии ц. н. с. областей коры, в которых регистрируется ВКО, было показано также А. А. Башкировым (1971).
Изучение ВКО в постнатальном онтогенезе у котят (Grafstein, 1959) показало, что вначале развивается отрицательная фаза ответа, а на 12-й день возникает положительная фаза, предшествующая у взрослого животного отрицательной. Автор считает, что положительная фаза обусловлена быстрыми проводящими волокнами, которые у новорожденных еще не миэлинизированы. Чисто отрицательный компонент ВКО у взрослого животного может возникнуть при слабых раздражениях, в то время как при тех же условиях погружение электрода в глубь коры приводит к регистрации положительной фазы (Grafstein, 1963). Из всех этих фактов было сделано предположение, что отрицательная и положительная фазы ВКО связаны с возбуждением различных групп каллозальных волокон.
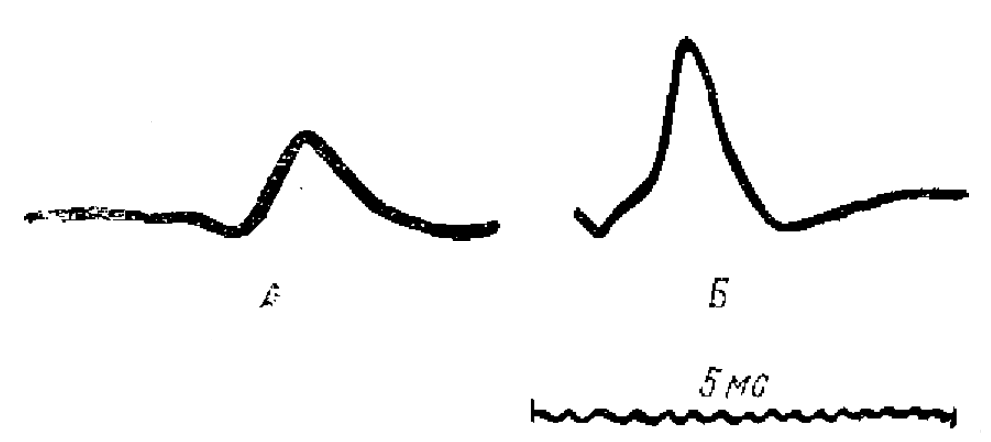
1 — начальное отрицательнее отклонение, 2—положительная фаза первичного отпета, 3 — отрицательная фаза первичного ответа, 4— положительная фаза вторичного ответа
Вторичные генерализованные ответы
Вторичные генерализованные ответы коры ретикуло-стволового происхождения. Мы уже дали общую характеристику ответу Форбса или вторичной генерализованной реакции ретикуло-стволового происхождения. Эта реакция возникает в обширных областях коры либо при одиночном электрическом раздражении стволовой части ретикулярной формации, либо при раздражении афферентного нерва. В последнем случае она возникает через определенный промежуток времени вслед за ПО. Отличается от ПО более длинным латентным периодом (30—80 мс) и возникает в обширных областях коры обоих полушарий (рис. 8). Лучше всего вторичная генерализованная реакция выражена в области передних латеральных извилин. Длинный латентный период этой реакции подразумевает, что она возникает благодаря распространению к коре импульсов по полисинаптическим путям. Старцль и др. (Starzl et al., 1951) уточнили пути распространения этой импульсации и пришли к выводу, что существуют два независимых пути такого распространения: внеталамический, через структуры суб- и гипоталамуса и внутреннюю капсулу к коре, и через таламус и его вентрально-медиальные ядра.
Как мы уже указывали, для генерализованной вторичной реакции ретикуло-стволового происхождения, хотя она и регистрируется широко в коре, существует ФМЛ (Purpura, 1955). Позже Пурпурой было показано, что при раздражении стволовой ретикулярной формации (от нижних бугров четверохолмия до заднего гипоталамуса) можно получить вторичную ретикуло-стволовую реакцию, проявляющуюся более локально во фронтальных и центральных областях коры и возникающую с более коротким латентным периодом (20—30 мс). Эти коротко-латентные ретикуло-стволовые реакции стали специальным предметом изучения ряда авторов (Абуладзе, 1968; Ильюченок и др., 1969, 1971; Лоскутова, 1970; Зиневич, 1969, и др.).
Таким образом, наряду с длиннолатентной генерализованной реакцией ретикуло-стволового происхождения в коре могут иметь место реакции ретикуло-стволового происхождения более коротколатентные и более локальные. Это показывает определенную специфику связен РФ с корой больших полушарий, однако функциональное их значение еще требует выяснения.
Вторичные генерализованные реакции ретикуло-таламического происхождения. Ритмическое раздражение неспецифических ядер таламуса приводит к возникновению в коре реакции вовлечения. Одиночное раздражение неспецифических ядер таламуса вызывает в коре одиночное колебание из комплекса реакции вовлечения. Этот ответ коры обычно регистрировался в виде большой поверхностно-отрицательной волны с большим латентным периодом (от 20 до 35 мс, см. рис. 9)(Arduini, Тегzuoio, 1951), В ряде случаев отрицательной волне предшествует незначительное по амплитуде положительное колебание, которое обычно связывали с активностью афферентных волокон, а отрицательную волну — с активностью корковых нейронов.
Специальные исследования Верцеано и др. (Verzeano et al., 1953) показали, что ответ коры на одиночное раздражение неспецифических ядер таламуса более сложен и состоит из нескольких фаз, где наиболее выражена отрицательная волна большой амплитуды, которой могут предшествовать 2 низкоамплитудные положительные волны. Вслед за этой большой отрицательной волной, определяемой как вторичная генерализованная реакция таламического происхождения, могли следовать еще несколько положительно-отрицательных колебаний низкой амплитуды.
Однако корковые ответы могли носить разный характер и, прежде всего, они разнились латентным периодом своих фаз. Авторы различают «быстрый» ответ, где латентный период высокоамплитудной отрицательной волны равен 26 мс, и «медленный» ответ с латентным периодом этой волны 41 мс. Эти ответы регистрировались в разных зонах коры: «быстрый» в передней сигмовидной, супрасильвиевой и цингулярных извилинах, «медленный» — в прореусе, орбитальной и задней супрасильвиевой извилинах.
Как показали Хэнбери и др. (Наnbегу et al., 1954) этот эффект зависел от раздражения разных ядер неспецифического таламуса: раздражение каудально расположенных ядер вызывало «медленный» ответ, а ядер, расположенных ближе к оральному полюсу таламуса — «быстрый» ответ (рис. 9).
Итак, мы рассмотрели группу ВП, объединенных под общим названием вторичные ответы коры больших полушарий. Безусловно, мы не охватили такого огромного материала, который накопился в литературе по вторичным ответам. Отсутствие единой терминологии и методических приемов исследования, трудности в трактовке полученных разными исследователями результатов пока не дают возможности более четкого представления об их генезе и функциональном значении.