Мы воспринимаем изменения во внутреннем и внешнем мире посредством рецепторов разных видов. Рецепторы можно подразделить на два больших класса экстерорецепторов (внешние) и интерорецепторов (внутренние). Экстерорецепторы позволяют получать информацию о событиях окружающего мира, а интерорецепторы — о том, что происходит в организме. Активность некоторых интерорецепторов никогда не достигает нашего сознания. Сенсорные элементы системы восприятия внешнего мира крайне специализированы. Существуют разные виды сенсорных модальностей, которые позволяют нам воспринимать изображения, звуки, движения тела, а также продуцируют боль, вкус, запах, осязание.
Важно отметить, что мы воспринимаем не объект как таковой, а импульсную активность выходных нейронов рецепторных органов. В свою очередь, эта активность нейронов возникает в результате взаимодействия рецептора и объекта. Области мозга, которые реагируют на стимуляцию определенных рецепторов, обычно рассматриваются как соответствующие сенсорные системы (зрительная, слуховая). Среди всех сенсорных систем наиболее изученной является зрительная. В настоящей главе мы уделим внимание рассмотрению зрительной, слуховой и соматосенсорной модальностей, поскольку данные системы доставляют в мозг наибольшее количество сенсорной информации и лучше всего исследованы.
Анатомия
Поля Бродмана и ядра таламуса
➥ Основная статья: Поля Бордмана
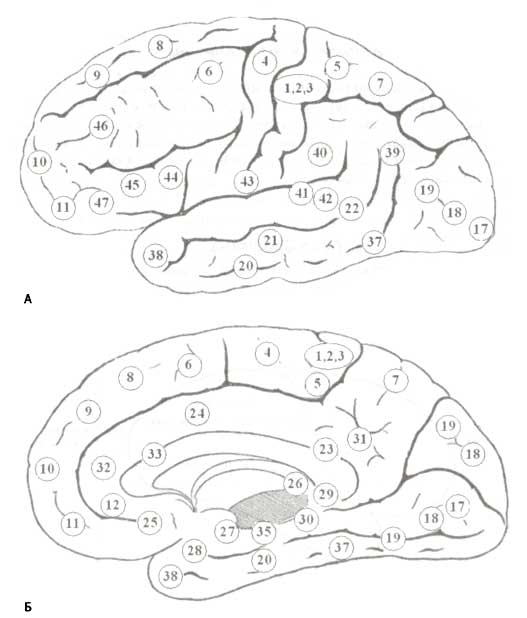
Схема человеческой коры с числами, соответствующими номерам полей Бродмана (определенных Бродманом в 1909 г. на основе цитоархитектонической организации коры).
Сенсорные сигналы от рецепторных органов достигают первичных сенсорных областей коры через релейные ядра таламуса. Основные сенсорные модальности локализуются в следующих областях: поле Бродмана 17 — первичная зрительная кора или область VI, ПБ 41 — первичная слуховая кора или область А1, ПБ 1 — первичная соматосенсорная кора или область S1. Латеральное коленчатое тело (ЛКТ) является релейным ядром для зрительной модальности сигналов, медиальное коленчатое тело (МКТ) — для слуховой, а заднее латеральное ядро — для соматосенсорной.
Топографическая организация
Каждая из первичных сенсорных областей коры характеризуется топографической организацией проекций. Нейроны первичной сенсорной коры и таламические ядра формируют сильно взаимосвязанную нейронную сеть. Важной особенностью этой сети является способность генерировать ритмическую активность в частотном диапазоне альфа-ритма. Альфа-ритм отражает неактивное состояние сенсорной системы — состояние, при котором сенсорная кора не участвует в активной обработке сенсорной информации, но за долю секунды может переключиться в активный режим.
Параллельные пути проведения информации

Сенсорные ядра таламуса (Б) выполняют воротную функцию, контролируя активность первичных сенсорных областей коры (А). На рисунке представлено три модальности — зрительная, слуховая и соматосенсорная. Схематическое изображение вентрального и дорсального зрительных проводящих путей (A). LGB — латеральное коленчатое тело, MGB — медиальное коленчатое тело, LP — латеральное заднее ядро таламуса.
В качестве примера рассмотрим пути проведения информации в зрительной системе. С механической точки зрения окружающий нас мир состоит из различных объектов различной локализации в пространстве, двигающихся с разной скоростью. Зрение обеспечивает нас полезным инструментом, позволяющим знать:
- что располагается в пространстве;
- где именно в этом пространстве это «что» располагается;
- как субъект может манипулировать этим «что» или сделать направленное движение в его сторону.
Для решения этих задач природа развила мозговые пути проведения информации (или потоки), передающие зрительную информацию разного вида. Эти пути берут свое начало от коленчатых тел таламуса, релейных ядер, состоящих из двух видов клеток, формирующих разные слои ядер: маленькие (парвоцеллюлярные) и крупные (магноцеллюлярные). Эти две системы (парвоцеллюлярная и магноцеллюлярная) остаются разделенными на уровне первичной зрительной коры (VI) и дают начало двум мозговым путям обработки зрительной информации.
Один из путей получил название вентрального, или «что»-системы (рис. 1) и включает вентролатеральную престриарную кору (части 18 и 19 ПБ) и нижневисочную кору. Стриарная, престриарная и нижневисочная кора составляют иерархически организованную систему, в которой сигналы передаются от первого уровня ко второму, от второго к третьему и так далее. Основной целью такой организации данного зрительного пути является разбиение первоначального образа на отдельные компоненты, характеризующиеся ориентацией, пространственной частотой и цветом.
Скорость и позиция зрительного объекта кодируется так называемой дорсальной системой обработки зрительной информации или дорсальным путем, включающим область теменной коры. Важной особенностью этого пути проведения зрительной информации является не только кодирование пространственных отношений зрительной сцены (т.е. «где» располагается объект), но и организация зрительно управляемых моторных программ (функция «как»). Задние области теменной коры имеют связи с премоторными центрами лобной доли, что позволяет обеспечивать реализацию целого класса ориентировочных и манипуляционных действий. Например, таких как фиксация взгляда на выбранном объекте, схватывание объекта и манипуляции с ним. В этом отношении дорсальная система тесно связана с процедурной памятью.
Вентральный и дорсальный пути обработки зрительной информации не являются строго параллельными, поскольку взаимодействуют на разных иерархических уровнях. Кроме того, в обеих системах существуют обратные связи, обеспечивающие возвращение информации от более высших уровней к низшим. Операции, связанные с вниманием и рабочей памятью, реализуются при помощи этих обратных связей (нисходящие потоки информации). В то же время связи между вентральной и дорсальной проводящими системами позволяют координировать обработку информации и формировать из различных сенсорных характеристик единый перцептивный образ.
Ядра подушки как координаторы информационных потоков. Обработка информации, осуществляемая в дорсальном и вентральном путях при участии престриарной, височной и теменной коры, не ограничивается внутренними связями между этими областями. Ядра подушки играют важную роль в координации процессов обработки зрительной информации (рис.1). Недавние исследования показывают, что нейроны подушки увеличивают активность при концентрации внимания на релевантном объекте и, напротив, подавляют свою активность в ответ на предъявление иррелевантного стимула. Тем самым обнаруженный эффект указывает на вовлечение этих нейронов в обеспечение контроля обработки зрительной информации в зрительной коре, что наиболее выражено при операциях, связанных с вниманием. Некоторые нейроны подушки, например, реагируют на движущиеся объекты в период фиксации взгляда, но не активны в период саккадических движений глаз, таким образом различая движение объекта от движений глаза. При этом участие ядер подушки в контроле процессов обработки информации соответствующими областями коры не ограничивается только зрительной модальностью: существуют данные, свидетельствующие о реакции нейронов подушки на соматосенсорные и слуховые стимулы.
После обработки сенсорная информация:
- перенаправляется в области префронтальной коры через последовательные (заднефронтальные) пути;
- распространяется к гиппокампу через ринальную кору.
Дополнительно существуют проекции «обратной связи» к дорсальной и вентральной системам обработки зрительной информации от префронтальной коры и гиппокампа.
U-образные кривые реактивности систем

Зависимость результирующей активности сенсорной системы от характеристик входа может быть представлена сигмоидальной функцией. При возрастании интенсивности сенсорного входа увеличивается и результирующая активность сенсорной системы. Однако это увеличение неоднородно и зависит от изначального состояния сенсорной системы. Система, активность которой увеличивается медленнее (быстрее) по сравнению с увеличением интенсивности входа, называется редуцирующей (усиливающей) сенсорной системой.
Общая активность сенсорной системы, как и любой нейронной сети, зависит от потока входящей информации и обычно описывается сигмоидальной функцией (рис. 2). В то же время реакция системы на предъявление одинаковых стимулов подчиняется инвертированному U-закону. Вкратце данный закон гласит, что реакция системы на стимул мала в тех ситуациях, когда ее общая активность или слишком мала (пороговый эффект), или излишне высока (эффект насыщения).
Увеличение и снижение активности сенсорных систем
Как будет вести себя сенсорная система при увеличении интенсивности стимуляции? Напрашивающийся ответ — увеличением своей активности (рис. 2). Однако характер этого увеличения будет зависеть от текущего функционального состояния системы. Если общий уровень активности и приток входящей сенсорной информации невелики (точка в нижней части кривой), то относительные изменения ответа системы будут больше относительных изменений сенсорного входа. Такие системы будут относиться к усиливающим (augmenting) сенсорным системам (рис. 2). В обратном случае, когда уровень общей активности системы высок, относительные изменения ответа будут меньше относительных изменений интенсивности стимуляции. Этот тип сенсорных систем можно охарактеризовать как редуцирующие сенсорные системы.
Например, в исследованиях на животных было обнаружено, что серотонинергические нейроны ствола мозга иннервируют слуховую кору. Такая иннервация, в свою очередь, приводит к зависимости общего уровня активности от уровня продукции серотонина. Хорошим индикатором функционального состояния слуховой системы является комплекс компонент ВП N1/P2. Так, если уровень серотонина и соответствующего притока входящей сенсорной информации увеличивается, то зависимость слуховых ВП от громкости снижается (на кривой рис. 2 изменение состояния системы отображается перемещением от левой точки на кривой направо). В исследованиях Джилланта (Gallinat et al., 2000) это свойство сенсорной системы и N1 /Р2 компоненты использовалось в качестве предиктора мощного ответа на ингибиторы обратного захвата серотонина при депрессии.
Диагностическая ценность сенсорных ВП
Негативность рассогласования
Одной из наиболее изученных компонент ПСС является негативность рассогласования. За последние 30 лет феномен HP интенсивно исследовался как с практической, так и с теоретической точки зрения. Сравнительно недавно HP стала использоваться для оценки фонологических и слуховых дисфункций при дислексии. В частности, в одном из таких исследований (Schulte-Korne et al., 1998) у двух групп подростков (здоровых и с диагнозом дислексии) проводился сравнительный анализ HP в ответ на речевые и неречевые стимулы. В качестве речевых стимулов использовались слоги (слог «да» — в качестве стандартного, «ба» — в качестве девиантного). Неречевыми стимулами были тоны (виды гармонической волны). Было показано, что амплитуда HP в ответ на тоны не отличалась между группами испытуемых. Однако при предъявлении слогов отмечалось относительное снижение амплитуды HP при дислексии по сравнению с HP здоровых испытуемых. Авторы интерпретировали полученные данные как отражение дисфункций в работе специфических фонологических систем, а не как генеральное нарушение процессов обработки слуховой информации. При этом когда дети, неспособные читать, тренировались с помощью зрительнослуховых упражнений, отмечалось увеличение амплитуды HP до нормальных значений.
Исследования шизофрении являются одним из интересных направлений клинических исследований с использованием HP. В частности, исследовалось влияние шизофрении на два типа генераторов HP, локализующихся во фронтальной и височной коре. Оказалось, что аффективное влияние заболевания больше выражено именно для фронтальных генераторов HP, чем для височных.
Исследования влияния процессов старения на амплитуду HP свидетельствуют о ее снижении с возрастом. Следовательно, как было показано в работах с использованием слуховых стимулов разной частоты, старение, по всей видимости, снижает объем слуховой памяти (не влияя на качество слухового восприятия в целом). Пациенты с диагнозом «болезнь Альцгеймера» при предъявлении стимулов с большим межстимульным интервалом (3 с) характеризуются сильным снижением амплитуды HP. HP используется и при прогнозах выхода из состояния комы. Наличие HP-подобной негативности в лобных областях при широком диапазоне изменений частоты стимуляции является предвестником возвращения сознательного состояния пациента.
Компонента сравнения
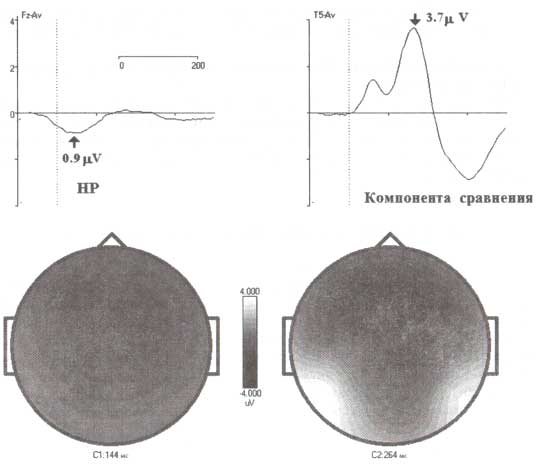
Негативность рассогласования (HP) рассчитывалась как разностная волна, получаемая при сравнении ВП при девиантном и стандартном тоне, предъявляемых в рамках oddball-парадигмы. Аналогично компонента сравнения рассчитывалась как разница между ВП для GO- и NOGO-стимулов, регистрировавшихся в двухстимульном GO/NOGO-тесте. Представлены усредненные по группе здоровых испытуемых (от 14 до 80 лет) ВП. Данные были взяты из нормативной HBI базы данных.
Даже короткий обзор литературных данных по практическому применению негативности рассогласования показывает, что с помощью HP можно определить психиатрические и неврологические отклонения от нормы. Однако величина эффекта в терминах амплитудных значений ВП достаточно мала, что ограничивает практическое применение HP в клинике. Незначительная величина эффекта частично может быть объяснена с позиций природы HP, средний уровень амплитуды которой 1 мкВ, что в 30—50 раз меньше фоновой ЭЭГ-активности.
С учетом этого более мощным диагностическим инструментом является компонента сравнения в зрительной GO/NOGO-парадигме. По крайней мере, амплитуда такой компоненты сравнения в 4 раза больше амплитуды HP (рис. 3). Более того, использование пространственной фильтрации с помощью метода независимых компонент позволяет дополнительно увеличить соотношение сигнала к шуму и обеспечить более эффективное применение в оценке индивидуальных ВП. Конечно, необходимы дальнейшие исследования в этом направлении, однако уже сейчас имеющиеся данные свидетельствуют о том, что данный метод является более мощным в сравнении с более старыми подходами к определению биологических маркеров (эндофенотипов) некоторых психических и неврологических расстройств.
Заключение
Мы оцениваем изменения во внутреннем и внешнем мире при помощи рецепторов разного типа. Существует несколько сенсорных модальностей, позволяющих воспринимать образы, звуки, движения тела и чувствовать боль, прикосновения, вкус, запах. Наша осведомленность о внешних или внутренних объектах не сводится к восприятию объектов как таковых, а заключается в обработке импульсной активности выходных нейронов рецепторных органов. Области мозга, нейроны которых отвечают на стимуляцию рецепторов конкретного типа, обычно рассматриваются как элементы соответствующих сенсорных систем (зрительной, слуховой системы и т.д.). В любой сенсорной модальности существует иерархическое подразделение на первичную, вторичную и ассоциативную кору. Ассоциативные области сенсорной коры отвечают за обработку наиболее сложных характеристик стимулов. Нейроны, кодирующие конкретные свойства сенсорного мира, формируют топографические организованные вычислительные карты. Например, нейроны, избирательная активность которых обеспечивает специфическую способность человека — восприятие лиц, локализуются в области височной коры и характеризуются специфическими компонентами скальпового сенсорного вызванного потенциала. Каждый класс объектов окружающего мира и каждый класс пространственных соотношений, как оказалось, обладает отдельной формой памяти. Категории памяти выступают в роли модели окружающей среды (Anticipating schemata). ВП, регистрируемые при активации этих моделей, могут быть разложены на несколько составляющих компонент. Мощным методом такого разложения ВП является недавно открытый метод независимых компонент. Выявляемые с помощью этого метода сенсорные компоненты характеризуются разной временной динамикой и пиковой латентностью, варьирующей от 110 мс в зрительной и от 80 мс в слуховой модальности. Разные компоненты генерируются в разных областях коры, начиная с первичных сенсорных, и распространяются вплоть до ассоциативных зон, включая переднюю поясную извилину. Психологические операции первичной зрительной (слуховой) обработки, сравнения (подразделяемые на детекцию физических и смысловых изменений) и вовлечения характеризуются разными компонентами. В зрительной модальности компоненты, отражающие операции сравнения и вовлечения, генерируются структурами вентрального и дорсального путей обработки зрительной информации. Операции сравнения в слуховой модальности, на иерархически низшем уровне, отображаются генерацией негативности рассогласования. HP отображает автоматический процесс оценки изменений, заключающийся в детекции рассогласования девиантного стимула со следом памяти, сформированным предшествующей повторяющейся слуховой стимуляцией. HP обнаружена в исследованиях, проводимых в рамках слуховой oddball -парадигмы, и используется в диагностических целях при разного рода психических и неврологических расстройствах.