Психология
Внимание как операция выбора
Представьте, что вы находитесь в комнате с механическими часами с закрытыми глазами. В тот момент, когда вы только что закрыли глаза, вы не слышите, как идут часы, но спустя некоторое время вы обращаете на них внимание. Внимание к звукам в некотором роде выглядит как «увеличение громкости» громкоговорителя: тиканье часов становится как бы громче. Переключение внимания на другой источник звука, например шепот вашего собеседника, сидящего в углу комнаты, способствует «выключению» звука часов. Именно так и работает наше внимание! Как видно из этого примера, внимание усиливает релевантную сенсорную информацию и подавляет нерелевантную. Эти операции усиления/подавления обозначаются термином «селективные операции» (операции выбора). Аналогично внимание как психологический процесс может быть определено как селективные операции в сенсорной модальности, целью которых является более тщательная (аккуратная) обработка релевантной сенсорной информации.
В начале XX века в мировой науке доминировала теория условных рефлексов Ивана Петровича Павлова и теория оперантного обучения Бурхуса Скиннера (Burhhus Skinner). В конце 1940-х и начале 1960-х в психологии было сформировано представление о различной природе глобальных состояний поведения, таких как возбуждение (активации) и сон. В исследованиях Джузеппе Моруззи (Giuseppi Moruzzi) и Хорасе Магуна (Horace Magoun) было показано, что стимуляция ретикулярной формации ствола мозга кошки вызывает пробуждение ото сна, тогда как стимуляция других, наоборот, индуцировала сон. В литературу был введен термин «пробуждение», обозначающий физиологическое состояние реактивности субъекта, и открыты его ЭЭГ-корреляты. В конце 1960-х новая научная парадигма, названная информационной обработкой, основанная на компьютерной метафоре, заменила бихевиоризм. Внимание стало одним из основных предметов этого направления исследований в психологии.
Сенсорные и моторные селективные операции
Селективные операции в более широком смысле не являются специфической характеристикой процесса обработки сенсорной информации, но в узком смысле представлены и на уровне моторных систем. Репертуар всевозможных моторных действий человека огромен. Мы можем танцевать, играть на музыкальных инструментах, говорить, читать и писать (некоторые из нас на нескольких языках)… Число элементарных действий, составляющих каждую из этих больших категорий действий, может быть также огромным. Просто представьте себе количество известных вам слов. Таким образом, в любой интервал времени доступно большое количество потенциальных действий. Целью нашего поведения является выбор действий, в том числе и тех, которые подходят наилучшим образом для текущего момента. В то же же время селективные операции не ограничены. Операции выбора выполняются и на когнитивном уровне — на уровне мыслей. Как говорил Вильям Джеймс, «думать — значит выбирать».
Подготовительная установка
В попытке объединения разных психологических сущностей в рамках единой концепции в классическом труде Эдварда Эвартса (Edvard Evarts), Ешиказу Шинода (Yoshikazu Shinoda) и Стивена Вайза (Steven Wise) «Neurophysiological approaches to higher brain functions», опубликованном в 1984 году, было введено понятие подготовительной установки. Под подготовительной установкой понимали состояние готовности к появлению определенного стимула или совершению движения. С учетом современных концепций можно говорить, что установочные реакции в сенсорной области связаны с вниманием, а в моторной или когнитивной — с исполнительными функциями.
Поведенческий паттерн в любой момент времени может быть разделен на две составляющие: исполнение когнитивно-моторного действия и его сенсорное восприятие. Естественно, сенсорные и моторные компоненты поведения взаимосвязаны. В действительности почти любое произвольное действие ассоциируется с восприятием того, как оно выполнено (восприятие напряжения мышц, позиции конечностей и т.д.) и как происходит взаимодействие с окружающим миром во время его реализации (восприятие изменения освещенности, акустических волн в окружающем воздухе и т.д.). И наоборот, почти любой перцептивный акт (оценка цвета, положения, конкретного объекта) связан с движением субъекта, его головы или глаз относительно внешнего мира (позиционирование взгляда в определенной точке видимого пространства как при слежении за летящим объектом, так и при рассматривании сложной фигуры).
С психологической точки зрения внимание, моторная подготовительная установка и селекция мыслей могут быть рассмотрены как похожие сущности, связанные с предварительной настройкой активности нейронных сетей. Несмотря на это, в условиях эксперимента такие элементы поведения исследуются отдельно. Исследования показали, что исполнительные и перцептивные функции реализуются разными нейронными механизмами. В обеспечении процессов выбора действий ключевую роль играют базальные ганглии в составе таламокортикальной системы. Селекция источников сенсорной информации обеспечивается за счет прямых и обратных связей разных кортикальных представительств сенсорных стимулов. Эти представительства имеют реципрокные связи с подкорковыми структурами (таламус, бугорки четверохолмия), что позволяет осуществлять операции по переключению внимания.
Обработка множества объектов
В повседневной жизни окружающий нас мир состоит из разнообразного множества загромождающих друг друга объектов разной формы, цвета, а также особенностей движения, локализованных в разных частях пространства. Наш опыт подсказывает нам, что мы не обрабатываем все окружающие объекты одновременно. В качестве демонстрации попытайтесь определить положение вертикальной серой полоски на рис. 1 (в верхней части слева). Как видно, на это требуется определенное время. Фокус вашего внимания последовательно перебирает несколько положений, пока не найдет нужный объект.
Здесь вполне уместна метафора «луч прожектора», когда мы рассматриваем процесс поиска цели. Это выглядит так, словно возможности нашей зрительной системы при обработке информации множества объектов в любой текущий момент времени ограниченны. На рис. 1 видно, как расположенные слева полоски разной ориентации и цвета конкурируют за внимание.
Операции вовлечения, отвлечения и переключения
Исходя из приведенного выше примера, внимание является динамическим процессом. В соответствии с мнением Михаэля Познера, мирового эксперта в психологии внимания, можно выделить три типа операций в этом динамическом процессе (рис 1 внизу). Во-первых, это операция вовлечения — усиление нейронного представительства источника информации, являющегося объектом внимания. Второй элемент внимания — операция отвлечения — заключается в подавлении источника информации, на который внимание не направлено. Третья операция — это операция перемещения или движения как промежуточное состояние при обработке двух разных источников информации. Упомянутые три типа реакций легко различимы на примерах, приведенных на рис. 1 (в середине), представляющих наложение визуальной сцены и траекторий движений глаз во время ее просмотра. Как можно заметить, во время поиска цели последовательные фиксации взгляда располагаются на разных полосках.
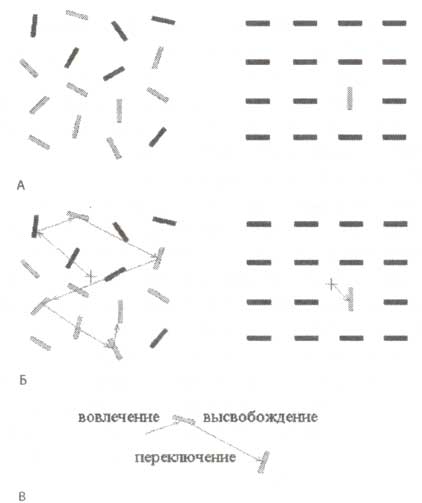
А. Изображения, иллюстрирующие два разных способа привлечения внимания. В соответствии с инструкцией внимание должно быть сконцентрировано на вертикальных полосках серого цвета. Поскольку возможности зрительной системы в обработке информации ограничены, то при одновременном предъявлении множества стимулов они будут конкурировать между собой. Очевидно, что процесс конкуренции сложен, если взглянуть на левое изображение (последовательный поиск цели), и относительно прост, если перевести взгляд на правое изображение (параллельный режим поиска цели). Следует отметить, что в первом случае нахождение цели обеспечивается в процессе поиска, контролируемого нисходящими влияниями от высших областей мозга, а во втором цель находится в результате управляемых стимулом восходящих процессов обработки информации. Б. Серыми стрелками обозначены перемещения внимания. Перемещения внимания могут быть явными и, соответственно, измеренными, например, при регистрации движений глаз, а именно — позиции центральной ямки глаза (фовеа) при рассматривании испытуемым изображения в поисках цели. Техника регистрации движений глаз во время рассматривания изображений впервые была использована отечественным ученым Александром Ярбусом. Перемещение внимания может быть также скрытным, т.е. осуществляться без видимых движений глаз. В. Три типа операций внимания, впервые описанные Майклом Познером (Posner et al., 1988): вовлечение, высвобождение и переключение.
Нисходящие и восходящие процессы
Важной предпосылкой существования внимания является наличие множества источников информации (например, несколько зрительных образов в зрительной модальности), которые конкурируют между собой. Ситуация борьбы разных стимулов за ресурсы может быть разрешена двумя способами: нисходящими или восходящими факторами влияния. Пример нисходящего фактора представлен в верхней левой части рисунка 1. Инструкция смотреть только на вертикальные серые полоски инициирует процессы высшего порядка, которые управляют саккадическими движениями глаз, определяющими перемещение внимание от одного стимула к другому, пока не будет найден искомый объект. Другой пример, приведенный на рис. 1 вверху справа, иллюстрирует действие восходящих факторов. Важным условием выраженности восходящих процессов является высокая степень отличимости целевого стимула по отношения к остальным (например, форма, цвет, специфическая ориентация). На рис. 1 (вверху справа) вертикальная серая полоска легко выделяется на фоне других стимулов и, соответственно, быстро и без усилий детектируется, обеспечивая ее высокую конкурентоспособность в борьбе за внимание. Этот девиантный стимул словно появляется на изображении. Таким образом, восприятие зрительных сцен происходит при взаимодействии нисходящих и восходящих влияний. Это взаимодействие составляет сущность внимания.
Анатомия
Сенсорные системы
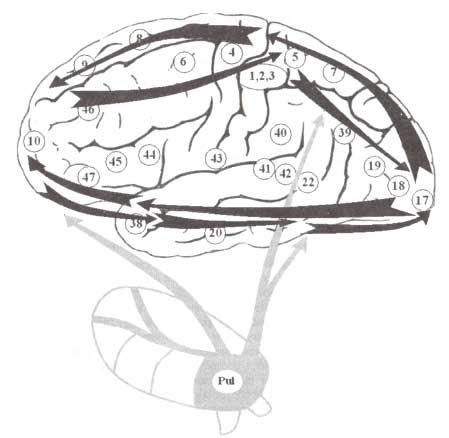
Внимание работает с сенсорной информацией и, следовательно, вовлекает элементы сенсорной системы (рис. 2). Помимо нисходящих и восходящих влияний на уровне иерархически организованных сенсорных областей мозга, обработка сенсорной информации контролируется подкорковыми мозговыми структурами — таламусом и верхними бугорками четверохолмия. Контроль информационных процессов в первичных и ассоциативных сенсорных областях коры осуществляется за счет их реципрокных связей с ядром подушки таламуса и ядрами ретикулярной формации, в соответствующей ее части. В обеспечение произвольных и непроизвольных перемещений взгляда вовлекаются верхние бугорки четверохолмия. Детальная обработка выбранных элементов окружающего трехмерного пространства контролируется ядрами подушки за счет реципрокных связей с сенсорными зонами коры.
Исполнительные системы
Функция внимания подразумевает реализацию операций выбора в сенсорной модальности. Однако сенсорное событие, включающее модуляцию внимания, не может быть отделено от действия. В частности, при переключении внимания с одной части визуальной сцены на другую должны быть выполнены также и действия, связанные с ориентацией глаз и расширением зрачка. Области, входящие в состав сенсорных систем, локализующиеся в задней части коры головного мозга, имеют реципрокные внутрикортикальные связи с передними областями исполнительных систем, вовлекаемых в процессы выбора действий, необходимых для реализации детальной обработки релевантной зрительной информации. Как уже упоминалось выше, различные механизмы выбора действий обеспечиваются фронтально распределенной исполнительной системой мозга.
Модуляция процессов обработки сенсорной информации
Взаимное подавление в исследованиях животных
Большинство наших представлений о нейронных коррелятах внимания было сформировано на основе исследований обезьян. Эти исследования чаще всего проводились в зрительной модальности. Схематический результат типичного исследования обезьян представлен на рис. 3 (для более подробного ознакомления см. Reynolds et al., 1999). Импульсная активность нейронов вентрального пути регистрировалась в ответ на предъявление зрительного стимула (например, полоска определенной ориентации) в рецептивном поле нейрона. В случае предъявления одного стимула, когда внимание было сфокусировано за пределами исследуемого рецептивного поля, регистрируемые ответы состояли из двух частей: ранний и поздний ответ. При одновременном предъявлении второго стимула в пределах этого же рецептивного поля наблюдалось подавление позднего ответа. Концентрация внимания обезьяны на первом из двух стимулов при игнорировании второго приводила к общему увеличение активности в ответ на такое сочетанное предъявление стимулов. Таким образом, полученные данные свидетельствуют о том, что одновременно предъявляемые одинаковые стимулы в пределах одного рецептивного поля, обрабатываются не независимо, а взаимодействуют друг с другом в режиме взаимного подавления. Как видно, внимание тормозит подавляющий эффект стимула, находящегося вне фокуса внимания, тем самым усиливая в ответ активность стимула, на который оно направлено. Коротко говоря, выявленные данные позволили считать, что внимание разрешает проблему противоборства среди множества стимулов, уравновешивая взаимное торможение между разными источниками информации. Это позволяет усилить обработку информации с тех источников, на которых сконцентрировано внимание, и параллельно ослабить влияние других.
Вовлечение подкорковых структур
Обработка сенсорной информации и модулирующее действие внимания на информационные потоки обеспечивается вовлечением двух подкорковых образований — ядер подушки таламуса и верхних бугорков. Верхние бугорки четверохолмия вовлекаются в генерацию саккадических движений глаз при произвольном поиске релевантных источников сенсорной информации. Их активность контролируется базальными ганглиями через окулярно-моторный проводящий путь. В ряде последних исследований приводится доказательство участия верхних бугорков четверохолмия не только в обеспечении целенаправленного поведения и саккад, но и в обеспечении скрытого внимания, осуществляемого при отсутствии наблюдаемых движений глаз.
Ядра подушки таламуса, благодаря реципрокным связям со структурами дорсального и вентрального путей обработки зрительной информации, а также другими сенсорными областями, играют важную роль в обеспечении внимания. За счет связей с областями префронтальной коры координируется работа исполнительных и сенсорных систем. При предъявлении стимула, на котором сконцентрировано внимание, обычно регистрируется увеличение активности нейронов подушки.

А. Рецептивные поля нейрона зрительной системы (обозначены квадратом) в трех разных ситуациях: слева—внутри рецептивного поля предъявлен только один зрительный стимул (горизонтальная полоска), а внимание сфокусировано вне рецептивного (обозначено серым цветом); посередине—в пределах рецептивного поля предъявлен дополнительный стимул (горизонтальная полоска), а внимание также направлено вне рецептивного поля; справа—внимание сфокусировано на горизонтальной полоске. Б. Типы реакцией нейронов на быстрое предъявление стимулов в трех разных ситуациях. Обращает на себя вниманието, что конкурирующие за внимание стимулы снижают позднюю компоненту ответа нейрона, которая восстанавливается при фокусировании внимания на одном из стимулов (в данном случае - горизонтальной полосе). В. Схематическое изображение сети нейронов, ответственной за обеспечение наблюдаемого эффекта. Пустыми кругами обозначены возбуждающие нейроны, а черными —тормозные нейроны. Слезет отметить, что внимание активируеттормозный нейрон, соответствующий первому стимулу, тем самым обеспечивая усиление детектирующих свойств нейронов, а также подавляя смежные нерелевантные нейроны. Обозначения: Ст. 1 — стимул 1; Ст. 2—стимул 2.
Негативные компоненты ВП, связанные с вниманием
Одной из первых парадигм исследования внимания с помощью метода вызванных потенциалов являлся тест дихотического прослушивания, разработанный Броадбентом в 1954 году. При одновременном предъявлении в правое и левое ухо стандартных и девиантных тонов (в рамках oddball-парадигмы) внимание испытуемого направлено на один из слуховых каналов. Сравнение ВП, зарегистрированных при предъявлении иррелевантных стимулов игнорируемого и целевого канала, характеризуется разностной компонентой — «разностной негативностью». Схематическое изображение получаемых результатов в такого рода исследованиях представлено на рис. 11.4 (слева). Более детальную информацию можно найти в книге Р. Наатанена «Внимание и функции мозга».
Результаты исследований, проводимых в другой парадигме, проиллюстрированы на рис. 4 (справа). Задачей испытуемых, которым в случайном порядке предъявлялись изображения шахматной доски (с красными и голубыми клетками), было попеременно обращать внимание на клетки одного или другого цвета. Разностный потенциал (при сравнении ВП стимула, на котором концентрировалось внимание, и ВП игнорируемого стимула) характеризовался негативной компонентой — «негативностью выбора». «Негативность выбора» считается ВП-показателем активности системы внимания на уровне вентрального пути обработки зрительной информации.

А. Процессная негативность в слуховой модальности; топограмма разностного ВП. полученного в результате сравнения ВП, зарегистрированных при стимуляции игнорируемого и прослушиваемого уха в условиях дихотического предъявления слуховых стимулов. Б. Селективная негативность в зрительной модальности; топограмма разностного ВП, полученного в результате сравнения ВП, зарегистрированных в ответ на предъявление изображения релевантного и игнорируемого цвета. Быстро и в случайном порядке в центре поля зрения предъявлялось изображение шахматной доски с клетками красного или синего цвета. В отдельных исследовательских сессиях внимание концентрировалось либо на синих, либо на красных клетках доски. Разностные ВП формировались при вычитании ВП на игнорируемый стимул из ВП на стимул релевантного цвета. В состав компонент, связанных вниманием, входит селективная негативность (см. Hillyard and Anllo-Vento, 1988).
Теменнофронтальная система в ПЭТ и фМРТ-исследованиях
Несмотря на то что настоящая статья посвящена контролю процессов обработки информации, осуществляемой мозговыми структурами задней части мозга, нельзя не упомянуть о взаимодействиях этих областей с фронтальными зонами мозга, традиционно связываемыми с рабочей памятью. В действительности понятия рабочей памяти и внимания тесно связаны: для того чтобы удерживать элемент в памяти, требуется внимание, и наоборот, внимание к определенным стимулам требует удержания в памяти ключевой информации о них. Кроме того, данные ПЭТ и фМРТ-исследований свидетельствуют об общности нейронных сетей, обеспечивающих эти операции. Традиционной находкой в разнообразных исследованиях со зрительно-пространственными заданиями, требующими затраты ресурсов внимания и рабочей памяти, являются активации в области теменных и лобных областей коры.
Нейрофизиология
Сенсорный дефицит и повреждения правой теменной коры
Как уже было сказано в предыдущей главе, нейроны теменной коры входят в состав дорсального пути обработки зрительной информации (системы «где»), ответственного за кодирование пространственного расположения объектов. Активность нейронов теменной коры ощутимо модулируется в задачах на пространственное внимание, когда внимание испытуемых концентрируется на разных участках окружающего пространства. Эти знания дают нам основания полагать, что повреждения теменной коры могут приводить к нарушениям пространственного внимания. Это подтверждается клиническими данными, в соответствии с которыми унилатеральное повреждение теменной коры приводит к сенсорному дефициту для объектов в контралатеральной части зрительного поля (относительно стороны повреждения). В более тяжелых случаях пациенты полностью игнорируют происходящее в определенной части зрительного поля, контралатеральной относительно повреждения. Например, такие больные могут читать только одну (правую или левую) часть книги, наносить макияж только на одну из половин лица или есть только с одной стороны тарелки. В менее тяжелых случаях сенсорный дефицит выражен меньше. В частности, дефицит может проявляться только в случаях предъявления конкурентных стимулов: как, например, в случае синдрома ослабления зрительного восприятия (visual extinction). Необходимо отметить, что пространственно-зрительный дефицит может быть вызван не только унилатеральным повреждением теменной коры, но и повреждениями в области лобной коры, передней поясной извилины, базальных ганглиев и ядер подушки таламуса (см. Kastner and Ungerleider, 2000).
Синдром Балинта
Полная утрата чувства пространства и способности выбора пространственных отношений между объектами наблюдается при билатеральном повреждении теменных долей. Поведенческие нарушения, вызываемые такими повреждениями, получили название синдрома Балинта. Пациенты с синдромом Балинта утрачивают способность воспринимать пространственную информацию (за пределами своего собственного тела) и с функциональной точки зрения слепы, за исключением сохранности перцепции одного зрительного объекта зрительной сцены. Они не могут ни определить положение видимого объекта, ни сказать, куда он движется (в их сторону или в другом направлении). Таким образом, полностью теряется пространственное восприятие.
Существуют аргументы в пользу того, что синдром Балинта — тип двойного сенсорного дефицита. Так, пациенты с сенсорным дефицитом могут не замечать как левую часть комнаты, так и левую сторону цветка. Однако при синдроме Балинта не замечаются обе части комнаты, но может целиком восприниматься единичный объект. Фактически такие бальные не видят ничего, кроме отдельных объектов. Таким образом, соотношение между двойным сенсорным дефицитом и синдромом Балинта не столь прямое, как может показаться на первый взгляд.
Нейрональные сети
Рекуррентная деполяризация апикальных дендритов
Повторяющаяся циклическая активность (как внутрикортикальная, так и таламокортикальная), по всей видимости, является важным условием обеспечения удержания внимания на определенном участке пространства (пространственное внимание) или объекте (непространственное внимание). Известно несколько моделей, имитирующих работу механизма внимания. Однако имеющихся эмпирических доказательств явно недостаточно для выбора наиболее адекватной модели. Вместе с тем один из элементов нейронной активности системы внимания наиболее, а именно поддерживаемая деполяризация апикальных дендритов пирамидных нейронов коры при различных установочных реакциях, часто подтверждается экспериментальными данными и входит в состав большинства моделей. Ключевую роль в обеспечении такой деполяризации играют возвратные связи, берущие начало от глубоких слоев областей коры высшего порядка с верхними слоями иерархически низших областей (рис. 11.5, на котором связи обозначены толстой стрелкой). Возвратные связи, активирующие по своей природе, вызывают деполяризацию апикальных слоев коры. По всей видимости, эта апикальная активность проявляется на ВП в виде медленной негативности при подготовительной установке. Апикальная деполяризация модулирует активность пирамидных клеток и снижает их порог активации. Пирамидные клетки средних слоев коры могут работать как детекторы совпадения, активируясь в случае деполяризации как апикальных, так и базальных дендритов.
Внимание и реакция активации
Из нашего опыта известно, что процессы сенсорного восприятия и распознавания объектов зависят от уровня общей активации. В состоянии бодрствования самому низкому уровню активации соответствует состояние дремоты. Противоположное экстремальное состояние — гипербдителъность (hypervigilance). В состоянии дремоты нам едва ли удается обратить на что-нибудь свое внимание, потому как пороги активации сенсорных нейронов настолько высоки, что даже появление значимого, релевантного стимула не вызывает их активации. Следовательно, работоспособность системы внимания крайне мала в таком состоянии. В состоянии гипервозбудимости наше внимание отвлекается различными нерелевантными стимулами, так как пороги срабатывания сенсорных нейронов крайне низки. В силу этого и в данном случае работоспособность мала. Таким образом, если попытаться определить способность сенсорной системы дискриминировать конкретные стимулы на фоне шума, приняв за «Р» работоспособность и за «А» общий уровень активации системы, то в результате получится так называемая инвертированная U-образная взаимосвязь.
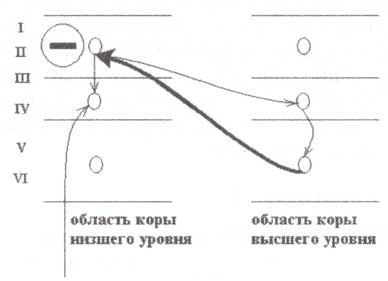
Слева направо — сети нейронов первичных и вторичных сенсорных областей. Контроль процессов внимания осуществляется за счет возвратного проводящего пути (обозначенных жирными линиями черного цвета) и деполяризации апикальных дендритов пирамидальных клеток (тонкие линии). Процессы деполяризации апикальных дендритов проявляются в виде негативности на регистрируемой со скальпа электрической активности.
Понятие возбудимости отражает фундаментальное свойство поведения и отчасти ассоциируется со способностью сенсорной системы к обработке информации. Складывается впечатление, что эффективные пороги активации сенсорных нейронов отражают способность мозга обрабатывать сенсорные стимулы (рис. 6). Установка этих порогов при общем уровне активации отлична от аналогичного механизма при подготовительной установке. В случае последней пороги выставляются весьма специфично, а действие такой настройки ограничено небольшим набором релевантных нейронов (относящихся к конкретным стимулам). Напротив, изменение общего уровня возбудимости приводит к общему «неспецифическому» изменению порогов. Эти изменения вызываются отдельной «неспецифической» системой.
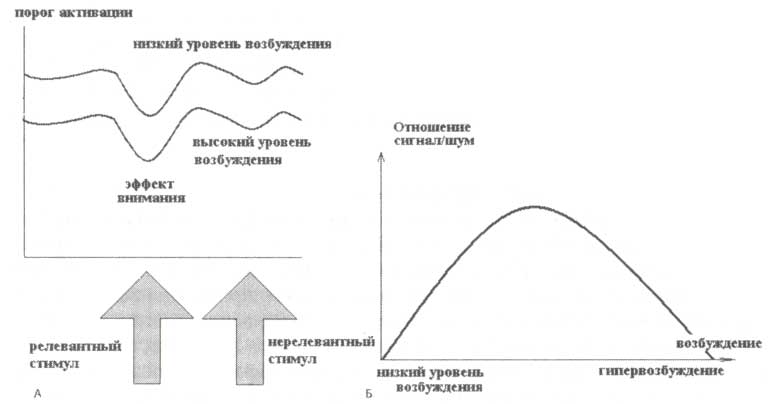
А. Пороги активации сенсорных нейронов: при высоком уровне бодрствования наблюдается неспецифическое снижение порогов как для релевантных, так и для нерелевантных стимулов; внимание же избирательно снижает пороги активации для релевантного стимула. Б. Слишком высокий или слишком низкий уровень бодрствования одинаково приводят к уменьшению соотношения сигнала к шуму.
Тонические и фазические реакции голубого пятна ствола мозга
Ключевым звеном системы неспецифической возбудимости являются ядра голубого пятна ствола мозга. Общий уровень активности нейронов голубого пятна строго коррелирует с изменением возбудимости в диапазоне состояний от глубокого сна до гипервозбудимости. Нейроны голубого пятна находятся в состоянии покоя в период сна, а именно фазы быстрых движений глаз (стадия парадоксального сна, или REM-стадия), и демонстрируют прогрессивную активацию при пробуждении. Фактически молчание нейронов голубого пятна является одним из физиологических параметров, которые используются для различения бодрствования и фазы парадоксального сна.
Нейроны голубого пятна показывают две разновидности активности: тоническую, или фоновую, активность, которая медленно изменяется вслед за медленными изменениями состояний мозга, и фазическую, которая отражает быстрый ответ системы на краткосрочную сенсорную стимуляцию. Считается, что фазическая компонента активности нейронов выполняет функцию временного усилителя (или фильтра), который интенсифицирует обработку наиболее значимых стимулов.
Ниже представлены свойства нейрона голубого пятна, доказывающие это мнение. Во-первых, латентность нейронных реакций, возникающих в ответ на стимул, весьма мала — около 100 мс. Во-вторых, только целевые, т.е. поведенчески значимые стимулы, вызывают активацию нейронов. В силу того что скорость проведения импульсов по немиеленизированным волокнам мала, активность нейронов голубого пятна может достигать коры в пределах 60—70 мс. Теоретически эти ответы могут усиливать поздние этапы обработки информации на уровне сенсорных систем.
Норэпинефрин как модулятор внимания
Нейроны голубого пятна являются единственным источником норэпинефрина в мозге. Норэпинефринергические нейроны немиелинизированы и, следовательно, обладают медленной проводимостью. Норэпинефрин может высвобождаться экстрасинаптически, производя нелокальный эффект. Эти свойства отражают тот факт, что эффект норэпинефрина на нейроны-мишени носит модуляторный характер. Как известно, модулирующий эффект регулятора зависит от рецепторов синапса. На уровне коры мозга существует два вида адренорецепторов — al и a2. При их активации генерируется возбуждающий постсинаптический потенциал, что приводит к снижению порога активации нейронов-мишеней Аксоны клеток голубого пятна иннервируют кору мозга неравномерно. Наибольшая плотность терминалей наблюдается в теменной, моторной и премоторной коре (рис. 7). Подкорковые структуры, принимающие участие в обеспечении внимания (подушка таламуса и верхние бугорки четверохолмия), также получают значительную часть входов от голубого пятна. Роль норэпинефрина в модуляции внимания объясняет существующий интерес к исследованию с помощью фармакологических подходов к изучению влияния препаратов на функции норэпинефрина. Один из недавно разработанных препаратов, атомоксетин, является блокатором обратного захвата норэпинефрина, что позволяет уменьшить симптоматику расстройства дефицита внимания.
Поздние позитивные компоненты вызванных потенциалов
➥ Основная статья: Вызванные потенциалы
Известно, что в интервале от 250 до 400 мс в ВП регистрируются позитивные компоненты, обозначающиеся как «Р300» или «Р3». Р3 генерируется при предъявлении значимых стимулов. Одна из компонент, принадлежащих к этой группе, Р3b, регистрируется в ответ на стимулы-мишени, т.е. стимулы, после которых следует моторное или когнитивное действие, например нажатие кнопки или подсчет количества стимулов-мишеней. Более ранняя компонента, называемый Р3а, увеличивается в амплитуде в ответ на неожиданное и заметное изменение в сенсорной стимуляции и отражает ориентировочную реакцию на подобное изменение стимула (рис. 7). Типичными экспериментальными условиями регистрации Р3а-компоненты является предъявление трех типов стимулов: стандартных, девиантных и редко появляющихся новых стимулов (например, собачий лай или телефонный звонок). Именно предъявление новых стимулов вызывает ориентировочную реакцию, переключение внимания и приводит к генерации Р3а-компоненты.
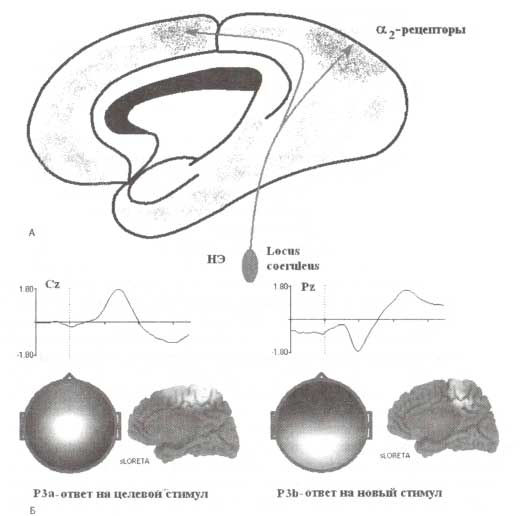
А. Голубое пятно (locus coereleus — LC) является источником норэпинефрина (НЭ) в коре. Аксоны нейронов голубого пятна обширно иннервируют кору головного мозга (не показано на рисунке). Наиболее плотно иннервируются теменные области, а также моторная и премоторная кора (обозначено скоплениями рассеянных точек серого цвета). Б. Р3а-компонента, генерируемая в тестовых условиях oddball-парадигмы стремя слуховыми стимулами: в типичном исследовании ВП в последовательности стандартных стимулов (тоны) редко предъявляются девиантные (целевые стимулы), перемежающиеся с редкими новыми отвлекающими стимулами (например, собачий лай). Испытуемые при предъявлении целевого стимула должны нажимать кнопку. Компонента Р3а генерируется при предъявлении отвлекающих новых стимулов. Целевые стимулы вызывают появление Р3b-компонент. Компоненты Р3а и Р3b характеризуются разной топографией и локализациями генераторов, вычисленной с помощью метода томографии низкого разрешения (sLORETA).
Р3b-компонента
Первое экспериментальное доказательство того, что компонента Р3b может отражать активность голубого пятна, было получено в работе Пинеды в 1989 году. В дополнение к этому существуют данные об общих характеристиках фазического ответа голубого пятна и Р3-компоненты: 1) обе реакции наблюдаются после предъявления стимулов-мишеней в рамках oddball-парадигмы; 2) обе реакции зависят от поведенческой значимости и внимания, привлекаемого к появляющимся стимулам; 3) наблюдается более выраженная активность в ситуации корректного ответа на стимул-мишень по сравнению с ошибочными ответами или пропусками в задачах на детекцию стимулов. С учетом вышесказанного была выдвинута гипотеза о том, что Р3b-компонента отражает фазическое увеличение притока входящей активности в кору, индуцируемое нейронами голубого пятна посредством продукции норэпинефрина (см. Jones and Cohen, 2005).
Проводимые нами в 1990-х годах исследования по изучению импульсной активности, регистрируемой с имплантированных электродов, выявили удивительный факт: Р3b-подобные компоненты могут быть обнаружены во множестве корковых и подкорковых структур (рис. 8). Паттерн мультиклеточной активности сильно зависел от локализации регистрирующего электрода. Так, области височной коры характеризовались активностью, сопоставимой с негативностью рассогласования, пиковая латентность которой варьировала в пределах 80— 120 мс и не проявляла зависимости от внимания. Компоненты, похожие на Р3b-компоненту, регистрировавшиеся в базальных ганглиях, ядрах таламуса и зонах лобно-затылочной коры, напротив, наблюдались только в активной модификации тестового задания, когда внимание испытуемого было сконцентрировано на детекции девиантных стимулов (требующих нажатия кнопки). Эти компоненты мультиклеточной активности по своим временным характеристикам совпадали со скальповой Р3b-компонентой. Однако по сравнению с присущей Р3b позитивностью регистрируемая активность была как положительной, так и отрицательной полярности. Более того, результаты исследования одного пациента показали, что характер Р3b-подобной активности существенно варьировал в зависимости от локализации электрода. По сути этот факт является свидетельством в пользу наличия нескольких разновидностей подобной активности, генерируемой в ответ на стимул-мишень, наблюдаемой в интервале 200—400 мс от момента его предъявления. Можно заключить, что скальповая Р3b-компонента в основном распространена над теменными областями коры. Однако при регистрации активности с имплантированных электродов картина меняется — выявляется широко распределенная мозговая система, объединяющая теменную, фронтальную кору, базальные ганглии и таламус.
В соответствии с литературными данными как в области клинических, так и теоретических исследований, компонента Р3b является наиболее изученной. Этому есть ряд причин. Во-первых, сама oddball-парадигма проста для применения практически в любых условиях исследования неврологических и психических больных. Во-вторых, Р3b — относительно большая компонента, которая легко выделяется на разностных ВП при сравнении ВП на девиантные стимулы мишени и игнорируемые стандартные стимулы. В-третьих, по всей видимости, Р3b характеризуется диагностической значимостью, убедительным доказательством которой являются значительные отклонения в величине этой компоненты при целом ряде дисфункций исполнительной системы, таких как шизофрения или синдром нарушения внимания с гиперактивностью.
Существует несколько предположений относительно функционального значения Р3b-компоненты. Наиболее популярная концепция, предложенная Дончиным в 1981 году, гласит, что Р3b отражает обновление рабочей памяти. То есть появление любого целевого стимула вызывает действие, после которого мозг обновляет содержимое рабочей памяти. В своей статье Дончин назвал этот процесс «обновление контекста». Концепция объясняла процессы на уровне психологии, но не предлагала конкретных нейрофизиологических или клеточных механизмов и в результате была раскритикована.
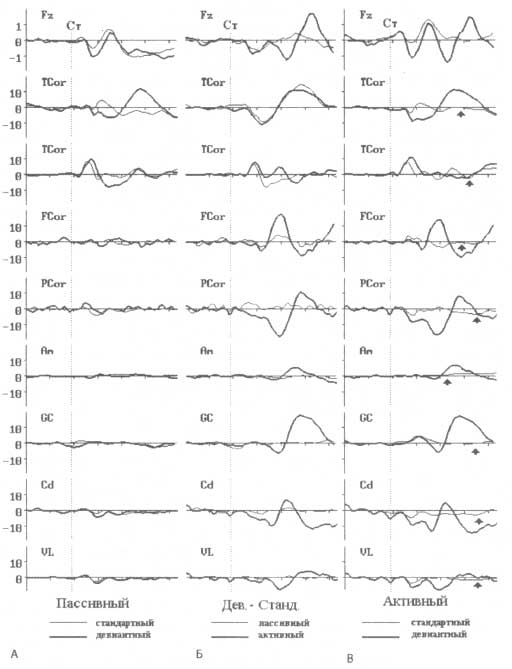
Вверху — скальповые ВП, зарегистрированные с Fz-отведения, усредненные по группе из 20 пациентов, которым по клиническим показаниям были имплантированы долгосрочные электроды. Амплитудная шкала — 1 мкВ. Ниже, сразу после верхнего ряда графиков ВП, представлены индивидуальные ВП, зарегистрированные с имплантированных электродов (амплитудная шкала—10 мкВ). ВП регистрировались во время выполнения акустического тестового задания в рамках oddball-парадигмы: стандартные стимулы (тоны длительностью 100 мс и частотой 1000 Гц) предъявлялись совместно с редкими и случайными девиантными стимулами (длительностью 100 мс и частотой 1300 Гц). Активный вариант теста (В) предполагал реакцию пациента в виде нажатия кнопки при предъявлении девиантного стимула. Пассивный вариант (Д) — чтение книги и игнорирование слуховой стимуляции. Графики разностных ВП в пассивной и активной модификации тестового задания представлены в среднем столбце (Б). Обозначения: ст.—стимул; дев.—девиантный; станд.—стандартный.
Р3а-компонента
Представьте, что вы управляете автомобилем, слушая музыку и наслаждаясь пейзажем за окном. Но неожиданное небольшое изменение в такте работы двигателя привлекает ваше внимание. Вы задаетесь вопросом: «Все ли в порядке?» Вы останавливаетесь, выходите из машины, открываете капот, смотрите и прислушиваетесь к двигателю. Данный пример демонстрирует то, что мозг создает сенсорную модель окружающего мира, удерживаемую в сенсорной системе. Когда происходит неожиданное изменение внешней среды, система обнаруживает это новое событие, что приводит к переключению внимания для его более тщательного изучения. Данный механизм позволяет людям (и животным) адаптироваться к постоянно меняющейся окружающей среде посредством так называемого ориентировочного ответа. Для того чтобы выделить бессознательную природу этого механизма, Иван Петрович Павлов называл его «ориентировочным рефлексом».
Существенный прорыв в понимании природы ориентировочного ответа был достигнут благодаря изучению скальповых ВП человека. Впервые компонента ВП, связанная с ориентировочной реакцией, а именно Р300а или Р3а, была описана в работе Саттона и др. в 1965 году. Несмотря на то что Р3а наиболее интенсивно исследовалась у людей, аналогичные компоненты регистрируются и у животных: макак, беличьих обезьян (саймири), кошек, собак, кроликов, дельфинов. Таким образом, Р3а может отражать мозговые процессы, свойственные млекопитающим.
В типичном для регистрации Р3а исследовании испытуемые выполняют тестовое задание по детекции стимула-мишени (тона), но при этом иногда среди нерелевантных стимулов случайным образом предъявляется новый неожиданный стимул (например, лай собаки). В отличие от стандартных стимулов и стимулов-мишеней появление нового стимула вызывает генерацию потенциала с пиковой латентностью около 200—300 мс (с момента предъявления начала стимула) и максимумом в центральных и лобных отведениях(рис. 7). Функциональное значение компонента Р3а и когнитивные процессы, связанные с ним, достаточно активно исследовались. В результате этих исследований стало возможным выявить четыре важных особенности Р3а. Во-первых, Р3а свойственно привыкание к последовательно предъявляемым новым стимулам, что выражается в снижении ее амплитуды в ситуации, когда эти стимулы становятся более ожидаемыми. Во-вторых, Р3а модально неспецифична — сопоставимые компоненты регистрируются при новых стимулах в зрительной, слуховой и соматосенсорной модальности. В-третьих, несмотря на то что обычно Р3а генерируется в ответ на сложные звуки, похожие потенциалы могут быть зарегистрированы и при предъявлении простых стимулов, отличающихся от текущего контекста. В-четвертых, Р3а регистрируется и в ответ на новый стимул в игнорируемом канале, что свидетельствует о выявлении нового стимула относительно фоновой стимуляции.
Следует отметить, что, как показывает sLORETA, нейрональные генераторы Р3а не локализуются исключительно на уровне премоторных областей. В нашем исследовании и работе, проведенной Эриком Халгреном (Eric Halgren) и др., Р3а-подобная активность встречается в различных корковых и подкорковых структурах: префронтальной, теменной, латеральной и медиальной височной коре, базальных ганглиях и таламусе (рис. 9). Очевидно, эта гетерогенная нейронная сеть объединяет несколько систем, выполняющих различные функции. Некоторые из этих структур, как, например, вторичные и ассоциативные области слуховой коры, ответственны за детекцию новизны посредством выявления характеристик стимуляции, которые весьма отличаются от фона. Другие (например, гиппокамп) вовлекаются в обеспечение кодирования контекстуально новых событий и их запоминание. И наконец, передняя поясная извилина и префронтальная кора играют важную роль в перераспределении внимания для более детальной обработки отклонений от фоновой стимуляции.
Наравне с фактом множественной генерации Р3а на сегодняшний день известно, что по крайней мере два нейромедиатора влияют на амплитуду Р3а: норадреналин (нейротрансмиттер, распространенный на уровне теменной и моторно-премоторных областей коры) и ацетилхолин, играющий важную роль в регуляции активности гиппокампа (связанной с процессами памяти).
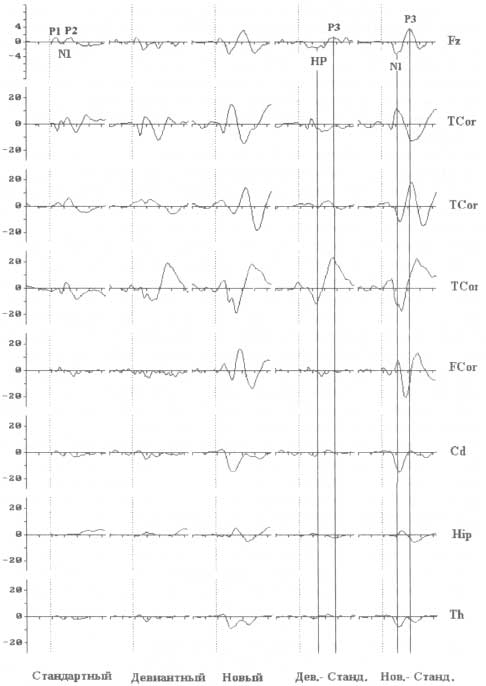
Вверху — скальповые ВП, зарегистрированные с Fz-отведения, усредненные по группе из 20 пациентов, которым по клиническим показаниям были имплантированы долгосрочные электроды. Амплитудная шкала — 4 мкВ. Ниже, сразу после верхнего ряда графиков ВП, представлены индивидуальные ВП, зарегистрированные с имплантированных электродов (амплитудная шкала — 20 мкВ). ВП регистрировались во время выполнения акустического тестового задания в рамках трехстимульной oddball-парадигмы: стандартные стимулы (тоны длительностью 100 мс и частотой 1000 Гц) предъявлялись совместно с редкими девиантными и новыми стимулами. Во время предъявления пациенты читали книгу. Графики разностных ВП, полученные при сравнении ВП на стандартные и девиантные, а также стандартные и новые стимулы представлены в двух правых столбцах. Обозначения: HP — негативность рассогласования; дев.—девиантный; станд. — стандартный; нов. — новый.
Диагностическая ценность Р3а и Р3b-компонент
Как считается, Р3а и Р3b компоненты отражают активность системы внимания, что обусловливает их широкое применение в диагностике расстройств, в которых предполагается нарушение функции внимания. Большинство исследований синдрома нарушения внимания с гиперактивностью свидетельствует о снижении амплитуды Р3b-компоненты у пациентов по сравнению с нормой.
Однако по сравнению с Р3b до сих пор не существует единого мнения о диагностической эффективности Р3а-компоненты. В некоторых исследованиях демонстрируется отсутствие значимой разницы между нормальными значениями Р3а и при СНВГ (например, см. Jonkman et al., 2000). Тем не менее существуют экспериментальные данные, согласно которым патологические изменения в генерации Р3а-компоненты (относительно нормы) могут отражать повышенную отвлекаемость детей с СНВГ (Gumenyuk et al., 2004).
Заключение
С физиологической точки зрения внимание ассоциируется с усилением обработки релевантной сенсорной информации и подавлением нерелевантной. Данные регистрации импульсной активности нейронов у животных указывают на то, что внимание решает проблему конкурирования стимулов за ресурсы с помощью механизма взаимного подавления репрезентаций множества одновременно представленных стимулов. В зрительной модальности пространственное внимание и внимание, относящееся непосредственно к объекту, реализуются на уровне дорсального и вентрального путей обработки информации соответственно. Нарушения пространственного внимания, называемые сенсорным дефицитом, ассоциируются с повреждениями в области правой теменной коры, структуры, входящей в состав дорсального зрительного информационного процессинга. Исследования ВП свидетельствуют о существовании ранних и поздних этапов обработки информации, которые по-разному модулируются процессами внимания. Ранние этапы характеризуются так называемыми негативностями выбора, а поздние — позитивными компонентами ВП: Р3а и Р3b. Механизм влияния внимания на генерацию ранних и поздних компонент ВП связывают с тонической или фазической активностью нейронов голубого пятна ствола мозга. Нейроны голубого пятна продуцируют регулирующий нейромедиатор норэпинефрин и обширно иннервируют кору головного мозга. На ранних этапах обработки информации тоническая активность этих нейронов играет важную роль в регуляции активности областей коры, влияя на генерацию ранних компонент ВП и внося существенный вклад в генерацию процессной негативности, регистрируемой на ВП. Атомоксетин, недавно открытый препарат, блокирует обратный захват норэпинефрина, снижая симптоматику нарушений внимания при СНВГ. Нейроны, продуцирующие норэпинефрин, характеризуются также сильно выраженными связями с теменной и премоторной областями коры, что обусловливает регуляцию поздних этапов процессов обработки информации. Считается, что фазическая модуляция активности коры этими нейронами отражается в компонентах Р3а и Р3b, которые широко используются как при исследовании здоровых испытуемых, так и в диагностических целях.