Дельта-ритм кортикальный — ЭЭГ-ритм, генерируемый при участии таламо-кортикального механизма. Преобладает в ЭЭГ, когда области коры отключены от соответствующих ядер таламуса.
Дельта-ритм таламический — ЭЭГ-ритм, регистрируемый с поверхности скальпа и генерируемый в таламусе при взаимодействии двух ионных токов таламокортикальных нейронов (ток катионов, деполяризующий мембранный потенциал, и кратковременный низкопороговый Са++-ток), ответственных за генерацию кальциевых спайков.
В ЭЭГ человека были выделены 2 типа дельта-колебаний: первый имеет корковое происхождение, в то время как второй генерируется в таламусе. Нейронные механизмы корковой генерации дельта-ритма неизвестны. Единственно, что мы знаем: хирургическое удаление таламуса, равно как и разъединения коры и таламуса, приводят к существенному повышению дельта-активности, регистрирующейся от скальпа. Эти факты указывают, что тип дельта-ритмов, генерируемый за счет внутрикорковых механизмов, вероятно, связан с некоторыми медленными процессами в пределах коры. Дельта-колебания могут быть связаны с медленно развивающейся синаптической депрессией. Мы знаем, что постсинаптическая депрессия может длиться сотни миллисекунд что соответствует дельта-диапазону частот. Положительные и отрицательные стадии дельта-колебаний, генерируемых в коре, могут быть связаны с синаптической депрессией, которая следует за текущей активацией.
Дельта-ритм сна

А. Таламокортикальная нейронная сеть. Возбуждающие нейроны и возбуждающие связи обозначены черным. Тормозные нейроны и тормозные связи обозначены серым. Б. Вверху - сдвиг от деполяризации к гиперполяризации, приводящий к переходу от режима отдельных спайков к режиму пачечной импульсации. В пачечном режиме потенциал мембраны колеблется с частотой дельта- активности, которая в свою очередь является результатом взаимодействия двух потоков ионов: катионного тока Ih, который активируется, когда нейрон гиперполяризован, и переходного низкопорогового тока Ca++-lt, который ответственен за возникновение Са*+-спайков. С. Открытие мембранных каналов в течение первых двух стадий возникновения Са++-спайков.
В отличие от дельта-активности, производимой в коре, механизмы таламической нейрональной генерации дельта-ритмов хорошо известны. При внутриклеточной регистрации активности в животных препаратах показано, что дельта-ритм может генерироваться отдельной таламокортикальной клеткой, то есть таламической клеткой, имеющей проекции к соответствующей корковой области. Схема таламокортикальной цепи, вовлеченной в генерацию дельта-ритма, изображена на рис. 1.
Здесь надо подчеркнуть, что та же цепь вовлечена в генерацию веретен сна (периодическая активность с частотой около 13 Гц) и альфа-ритмов (периодическая активность с частотой около 10 Гц) во время бодрствования. Эти ритмические активности проявляются на различных уровнях поляризации таламокорковых клеток: альфа-ритмы появляются, когда таламокорковые нейроны относительно деполяризованы, сонные веретена — когда эти клетки относительно гиперполяризованы, дельта-ритмы появляются на самом глубоком уровне гиперполяризации таламокорковых нейронов.
Для генерации веретен сна и альфа-ритмов необходимы свойства сетей, такие как синхронизированное торможение ретикулярными нейронами таламуса. В отличие от сонных веретен и альфа-ритмов дельта-волны генерируются отдельными клетками. Ритм генерируется взаимодействием двух токов (и следовательно — двумя типами ионных каналов) в таламо-кортикальных нейронах (рис. 2).
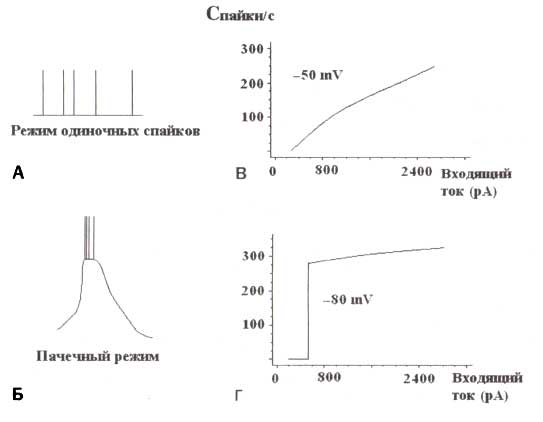
Схематическое представление тонического (А) и пачечного (Б) режима импульсаций таламокортикальных клеток. В, Г. Схематическое представление отношений ввода — вывода для отдельной клетки в этих двух режимах. Входная переменная — амплитуда деполяризующего импульса тока. Результирующая переменная — частота разрядов клетки. Исходные потенциалы 50 мВ для тонического режима и -80 мВ для пачечного режима.
Первый ток называют Ih — это катионный ток, который активизирован, когда нейрон гиперполяризован, и, как следствие, он возвращает мембранный потенциал из гиперполяризованного состояния к менее поляризованному. Второй поток называют It — это низкопороговый ток кальция. Во время деполяризованного состояния нейрона (состояние бодрствования) этот ток отсутствует. Он появляется только при гиперполяризации нейрона, во время глубокого сна из-за подавления сигналов от восходящей активационной системы ствола мозга, так же как из-за подавления входов от других мозговых систем.
It поток ответственен за генерацию так называемых Са++-спайков. Са++-спайк представляет собой ток ионов Са++ внутрь, который деполяризует мембрану »иламонор1иклльиых нейронов. I, управляется двумя воротами: активационными и инактивационными, It активируется при – 70 мН, при относительно низком порога, поэтому он называется низкопороговым и приводит к возникновению низкопорогового кальционного спайка. В этом его отличие oт обычного Na+ спайка, который имеет более высокий порог активации, соответствующий приблизительно 50 мН. Другого отличия между обычными Nа’-спайками и низкопороговыми Са++-спайками — продолжительность спайков.
It-поток отвечает за возникновение Са++-спайков. На вершине фазы деполяризации Са++-спайка нейрон генерирует вспышку обычных K +-Na+-спайков. Эти спайки через таламокорковые пути передаются соответствующим корковым областям и генерируют постсинаптические потенциалы, которые, в свою очередь, регистрируются от скальпа в форме дельта-ритма.
Генерация низкопороговых кальциевых спайков посредством It-тока впервые была продемонстрирована в таламических нейронах Rodolfo Llinas и Jahnsen в 1984 г., а также была зарегистрирована in vivo Marcelo Steriade и коллегами. Безусловно, можно отметить, что большое влияние на наше понимание клеточных механизмов генерации ЭЭГ было оказано McCormic и его сотрудниками.
Низкопороговый пачечный режим пульсаций таламических нейронов
Дельта-колебания связаны с так называемым пачечным режимом пульсаций таламо-кортикальных нейронов, который появляется только тогда, когда эти нейроны сильно гиперполяризованы. В этом состоянии отношения между входным и результирующим импульсом отдельной клетки представляют собой ступенчатообразную функцию, отражающую тот факт, что выходная частота разрядов таламо-кортикальных нейронов не зависит от входящего тока (внизу рис. 2). Этот режим пульсаций нейронов отличается от тонического состояния (или единичных разрядов), в котором таламо-кортикальные нейроны отвечают на входящие деполяризационные токи линейной зависимостью: чем больше входящий ток, тем больше разрядов на выходе.
Кальциевые спайки отдачи — не единственный способ вызвать пачечный режим разрядов. В коре существует целый особый набор нейронов, которые генерирую спайки в пачечном режиме. Этот режим может определяться внутренними особенностями нейронов или нейронной сети, к которой принадлежит нейрон. Пачечный режим пульсаций корковых клеток, вызванный деполяризующими токами, отличается от состояния пачечного режима пульсаций таламических нейронов (релейных и ретикулярных), вызванного гиперполяризующими токами.
Патологические ритмы дельты
Феноменология дельта-активности, появляющаяся при некоторых серьезных мозговых отклонениях, очень сложна. В главе известной книги Electroencephalography под редакцией Ernst Niedermeyer и Fernando Lopes Da Silva, названной Nonspecific abnormal EEG patterns, Frank Sharbrough суммировал все известные феномены, связанные с дельта-активностью в несколько категорий. Эти категории: неустойчивая ритмичная дельта-активность (IRDA), лобная неустойчивая ритмичная дельта-активность (FIRDA), постоянная неритмичная дельта-активность (PNDA). Например, центральную PNDA рассматривают как один из самых надежных признаков серьезного локального мозгового нарушения типа инсульта или мозговой травмы. Необходимо подчеркнуть, что ЭЭГ отражает главным образом корковые явления, в то время как подкорковые структуры имеют очень небольшое отражение в ЭЭГ. Таким образом, обнаруженные зоны повреждения мозга по данным ЭЭГ и трехмерной локализации пораженного участка мозга, например, с использованием возможностей компьютерной и магнитно-резонансной томографии, не обязательно будут совпадать. Более того, иногда видимые на ЭЭГ нарушения не картируются более высокотехнологичными КТ- и МРТ-методами и наоборот существенные нарушения по данным перечисленных методик в ЭЭГ могут характеризоваться незначительными изменениями. Для того чтобы иметь объективное представление о нарушениях в мозге человека при использовании метода ЭЭГ, необходимо визуальный анализ ЭЭГ подкреплять данными спектрального анализа и результатами исследований вызванных потенциалов.