Р3b как компонента, отражающая операции вовлечения
Компонента Р3b, или, точнее говоря, Р3b-компоненты, подробно рассматривались в предыдущей главе, посвященной описанию компонент ВП, характеризующих работу системы внимания. Несмотря на то что наибольшая плотность генераторов Р3b локализуется в области теменной коры, что и позволяет связывать данную компоненту с системой внимания, в генерацию этой волны вовлечена нейронная сеть, объединяющая фронтальные, височные, теменные области коры и базальные ганглии (таламокортикальными взаимодействиями).
Сенсорное сравнение
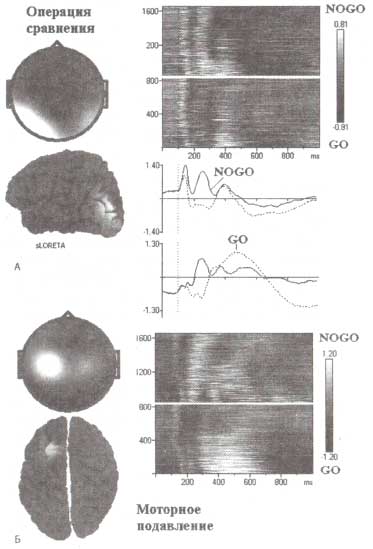
Метод независимых компонент применялся при анализе ВП, зарегистрированных в ответ на GO- и NOGO-стимулы, предъявлявшиеся в рамках двухстимульного GO/NOGO-теста. ВП-данные здоровых испытуемых возрастом от 1 до 89 лет были взяты из нормативной HBI базы данных. А. Компонента сравнения. Б. Компонента моторного подавления. Вертикально уложенные тонкие горизонтальные окрашенные полосы (цветом обозначается амплитуда), каждая из которых представляет изменение во времени амплитуды соответствующей компоненты ВП отдельного испытуемого. Представлены GO- и NOGO-условия. "Голограммы соответствующих компонент, локализация генераторов компонент, вычисленная с помощью метода томографии низкого разрешения (sLORETA), а также временная динамика ВП представлены в виде отдельных графиков для GO- и NOGO-стимулов.
В нашей повседневной жизни мы всегда стараемся «смотреть вперед» и заняты подготовкой к реализации предстоящих действий. Однако контекст окружающей действительности быстро меняется, что иногда требует от нас подавления ранее планируемого действия. Подавление подготовленных действий обеспечивается в результате сложных мозговых процессов, одним из элементов которых является активность латеральной префронтальной коры. Как уже было сказано выше, префронтальная кора получает информацию от сенсорных систем (зрительной, слуховой и соматосенсорной), на основе которой и принимает решение в пользу действия (GO) или подавления (NOGO). Далее возможна следующая спекуляция: для осуществления выбора действия мозгу в первую очередь необходимо выполнить сравнение текущей сенсорной ситуации с сенсорной моделью и выявить в случае наличия рассогласование. Операции сравнения сенсорных сигналов протекают на уровне сенсорных систем, располагающихся в задней части мозга. Продолжая рассуждения, логично предположить, что результаты таких операций сравнений передаются в префронтальную кору и активируют мозговые процессы, ответственные за подавление подготовленных действий.
Примечательно, что данные предположения получают экспериментальное подтверждение. Так, на рис. 1 продемонстрирован результат анализа ВП с применением метода независимых компонент, полученных в исследовании, проводимом в рамках GO/NOGO-парадигмы. Как видно из рис. 1, NOGO- и GO-стимулы вызывают генерацию разных независимых компонент ВП, генерируемых в височной и премоторной коре. На уровне височной коры NOGO-стимул характеризуется дополнительной компонентой (в сравнении с GO-пробой). Однако в области левой премоторной коры предъявление этих стимулов приводит к прямо противоположным реакциям. Разный характер временной динамики компонент свидетельствует о наличии функциональных отличий между ними. Вполне вероятно, что первая компонента, характеризующаяся позитивностью в височной области, может отражать операцию сравнения сенсорных сигналов — детекцию изменения.
Подавление моторной активности
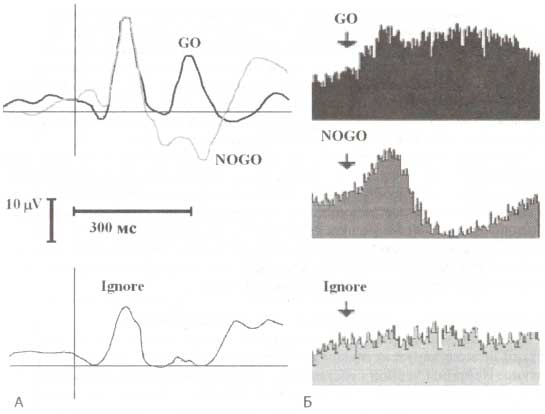
А. Интракраниальные ВП. Б. Ответы таламических нейронов на GO-, NOGO- и игнорируемые стимулы трехстимульной oddball-парадигмы. (А): по оси Y — усредненные потенциалы, которые были зарегистрированы с электродов, имплантированных в вентральную область таламуса в ответ на GO- (черная линия), NOGO- (серая линия) и игнорируемые (Ignore, внизу) стимулы. (Б): по оси Y — усредненная импульсная активность нейронов, зарегистрированная с тех же электродов. По оси X — время (неопубликованные данные из архивов лаборатории).
Можно также предположить, что вторая компонента (рис. 1) связана с моторным подавлением. Если сравнить эту компоненту с паттерном вызванной активности таламуса (рис. 2), то обнаруживается удивительное сходство между скальповым ВП и локальным потенциалом на таламическом уровне. Факт этого сходства, а также локализация генераторов компоненты является подтверждением наших предположений. Следует оговориться, что компонента, связанная с моторным подавлением во фронтальных отведениях, характеризуется негативными значениями амплитуды.
Нейронные механизмы подавления моторного ответа, как было показано выше, вовлекают процессы, происходящие в базальных ганглиях. Особенностью организации связей коры мозга является наличие длинных возбуждающих внутрикортикальных связей и отсутствие таковых для тормозных влияний, что обусловливает исключительно локальный характер процессов торможения, инициируемых структурами коры. Это означает, что сама кора не в состоянии полностью подавить программу действий. Наиболее вероятный механизм подавления подготовленных действий подразумевает вовлечение таламокортикальных взаимодействий, контролируемых базальными ганглиями посредством непрямых проводящих путей, с важным звеном системы, локализующимся в премоторной коре.
Подавление действия
Одним из примечательных компонент ВП, получаемых в результате анализа независимых компонент ВП в рамках парадигмы GO/NOGO, является третья NOGO-компонента (рис. 1). Пространственное распределение, временная динамика данной компоненты, а также результат прямого сравнения с ВП при GO-стимулах существенно отличаются от всех остальных компонент. Кроме того, NOGO-компонента является симметричной, характеризуется единственной выраженной пиковой латентностью в 340 мс, а ее генераторы широко распределены на уровне премоторной и моторной коры. Важно, что этот эффект наблюдается только в случае предъявления NOGO-стимулов. Также существует зависимость показателей компоненты от возраста: с возрастом отмечается снижение амплитуды и увеличение латентности. При этом амплитуда NOGO-компоненты негативна во фронтальных областях. Для различения асимметричной компоненты, связанной с моторным подавлением, данную компоненту назвали «компонентой подавления действия». Ситуация такова, неизвестно, почему мозг генерирует эту компоненту после продукции специфической компоненты подавления моторной реакции. Неизвестно, генерируется ли данная компонента посредством ганглионарноталамокортикальных взаимодействий или за счет активности самой коры. Но то, что действительно понятно, — что в терминах амплитуды компонента подавления действий в три раза превосходит компоненту моторного подавления и, следовательно, вполне вероятно, обладает лучшей диагностической ценностью (в сравнении с компонентой моторного подавления).
Регистрация активности с имплантированных электродов
Проводя лечение различных больных с помощью имплантированных электродов, можно регистрировать внутримозговые слуховые ВП в условиях двухстимульной исследовательской GO/NOGO-парадигмы. Электроды имплантировались в разные корковые и подкорковые структуры исходя из диагностических и терапевтических целей. Пациентам, страдающим паркинсонизмом, электроды имплантировались в область таламуса и базальных ганглиев. Другие области-мишени применялись при лечении эпилепсии — медиальная (включая гиппокамп) и латеральная височная кора. При обсессивно-компульсивном расстройстве электроды локализовались в области передней поясной извилины и смежных областях префронтальной коры.
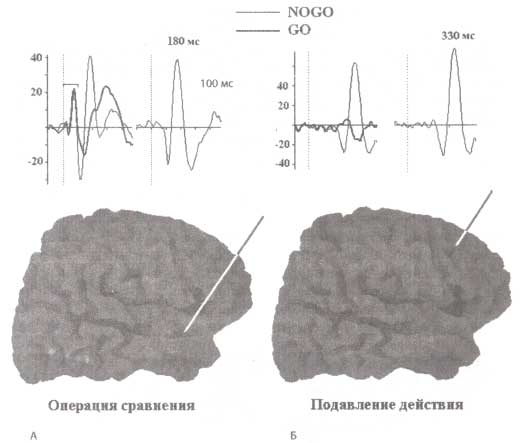
Элетроды были имплантированы пациенту по терапевтическим показаниям (для проведения лечения и диагностики). Пациент выполнял двухстимульный GO/NOGO-тест. Электроды были локализованы в ассоциативной слуховой коре (А) и дорсолатеральной префронтальной коре (Б). Регистрировались ВП для GO, NOGO и GO/NOGO (разность) условий. Внизу рисунка проиллюстрированы области имплантации электродов.
На рис. 3 представлены ВП, зарегистрированные с двух электродов, располагавшихся в височной доле и префронтальной коре. Пациенты выполняли тестовое задание — модифицированную версию двухстимульного теста GO/NOGO-парадигмы. Как видно из рис. 3, предъявление NOGO-стимула на уровне височной коры характеризовалось негативно-позитивной компонентой с соответствующими пиковыми латентностями 100 и 200 мс. В префронтальной коре наблюдался сопоставимый паттерн активности в ответ на NOGO стимул, но с большими значениями ЛП, а именно на 150 мс позже своего височного аналога. Следует также отметить и наличие ранней компоненты с латентным периодом 70 мс, зарегистрированного с височного электрода. Впрочем, данная ранняя компонента была одинакова для GO- и NOGO-стимулов и, по всей видимости, отражает ранние этапы сенсорной обработки стимулов. ВП префронтальных отведений, напротив, характеризовались только поздними компонентами и только при предъявлении NOGO-стимулов. Если проводить сравнение внутримозговых и скальповых ВП, то можно говорить о том, что в конкретном рассматриваемом случае ВП, зарегистрированные с височной коры, отражают операцию сравнения. В то же время локальный потенциал префронтальной коры ассоциируется с операциями подавления действий.
В изучении процессов мозга человека, связанных с подавлением действий, кроме GO/NOGO-парадигмы также применяется так называемая задача стоп-сигнала. Аналогично GO/NOGO-парадигме в задачах со стоп-сигналами выполнение текущей деятельности должно быть прекращено по предъявлении соответствующего сигнального стимула. Полученные за последние время экспериментальные данные (в рамках обоих исследовательских подходов) свидетельствуют в пользу того, что ключевая роль в подавлении действий принад лежит вентролатеральной префронтальной коре. В онтогенезе данная область развивается одной из последних и именно поэтому неполное развитие этой части префронтальной коры может рассматриваться как возможная причина синдрома импульсивности, по крайней мере у некоторых подгрупп детей с СНВГ. Так, в работах Адама Арона из Кембриджского университета (Англия) была показана корреляция выраженности повреждения правой вентролатеральной префронтальной коры (но никакой другой области) и показателей процессов подавления действий (время реакции при предъявлении стоп-сигналов): при более обширных повреждениях наблюдалось большее замедление времени реакции.
N200 как компонента моторного подавления
Исследования нейронных механизмов подавления моторной деятельности берут начало в 70-х годах прошлого столетия. Наиболее важные открытия были сделаны японскими исследователями Гемба и Сасаки. Ими были выявлены специфические механизмы моторного подавления на уровне моторной и премоторной коры. Так, в исследованиях на обезьянах было показано, что возбуждение нейронов центральной извилины (principle sulcus) на фоне регулярных ответов может приводить к снижению активности премоторной и моторной коры и даже задержке или полному подавлению ответа. При этом прямая электрическая стимуляция нейронов этой области вызывает подавление ответа на GO-стимул. В исследованиях человека нейронный механизм процесса подавления изучался с помощью метода ВП (скальповые). Во множестве таких исследований регистрировалась негативная компонента N200, распределенная в лобных областях и достигающая максимальных значений спустя 200—260 мс после предъявления NOGO-стимула. В сравнении с GO-стимулом N200 при NOGO характеризуется большей амплитудой и считается индикатором процессов подавления в парадигме GO/NOGO. Сопоставимая N2-компонента наблюдается и при предъявлении стоп-сигналов. Однако следует отметить, что во всех классических работах N2-компонента получается в результате вычисления разностной волны между ВП для GO- и NOGO-стимулов.
Ниже плказано, что классическая N2-компонента также наблюдается и при анализе полученных данных в виде фронтально распределенной негативности, регистрируемой в ответ на NOGO-стимул. На рис. 4 представлен ВП, усредненный по той же группе пациентов (исследовавшийся в рамках GO/NOGO-парадигмы), что и ВП. Как видно, NOGO-стимул характеризуется выраженным негативным отклонением с ЛП 260 мс, который может считаться аналогом классической N2-компоненты. Негативная компонента сменяется позитивной, также характеризующейся фронтальной топографией. Если провести сравнение ВП на рис. 4 и трех независимых компонент, полученных при анализе ВП методом независимых компонент (рис. 1 и 5), то можно заметить одну особенность: негативная компонента традиционного ВП в действительности является суммой этих трех разных независимых компонент.
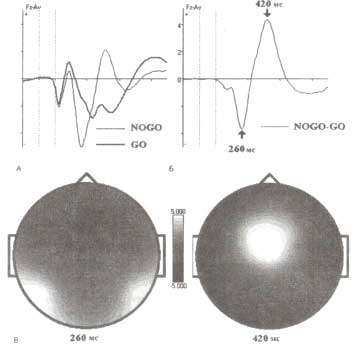
Представлены усредненные по группе испытуемых ВП (испытуемые от 1 до 89 лет), рассчитанные на основе ВП-данных из нормативной HBI базы данных. А. ВП для GO- и NOGO-стимулов двухстимульного GO/NOGO-теста в Fz-отведении. Б. Разностный потенциал (NOGO-GO). В. Топографии разностных ВП, вычисленных для двух событий — операции сравнения и подавления.
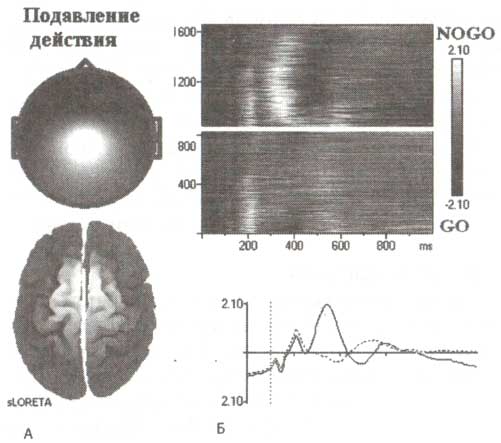
Метод независимых компонент применялся при анализе ВП, зарегистрированных в ответ на GO- и NOGO-стимулы, предъявлявшихся в рамках двухстимульного GO/ NOGO-теста. ВП-данные здоровых испытуемых возрастом от 7 до 89 лет были взяты из нормативной HBI базы данных. А. Топография и локализация генераторов компоненты подавления, вычисленная с помощью метода томографии низкого разрешения (sLDRETA). Вертикально уложенные тонкие горизонтальные окрашенные полосы (цветом обозначается амплитуда), каждая из которых представляет изменение во времени амплитуды соответствующей компоненты индивидуального ВП. Б. Временная динамика ВП представлена в правой нижней части рисунка.