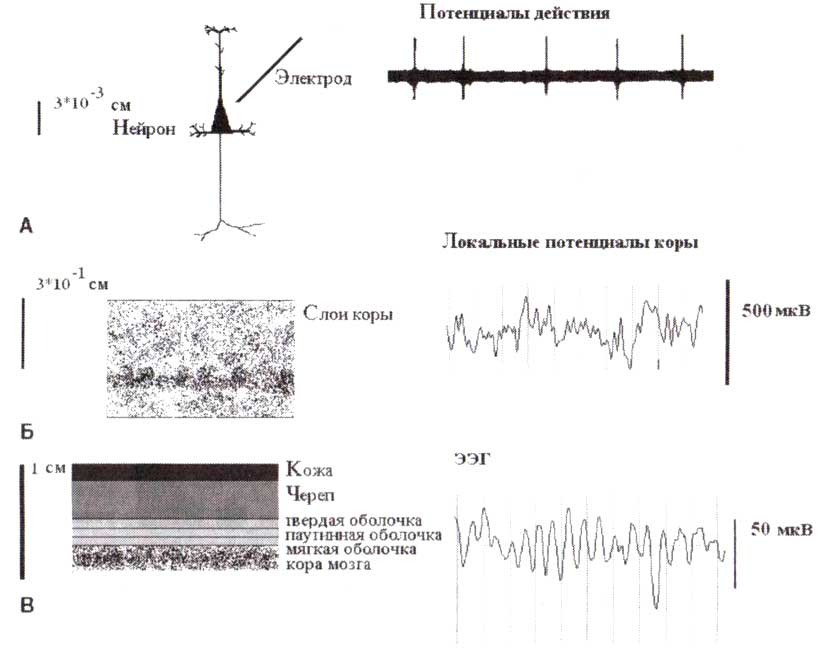
Уровень одиночного нейрона соответствует 30 микронам пространственной шкалы и длительности спайка, равной 1 мс. Локальные потенциалы измеряются с помощью макроэлектродов, помещенных в ткань мозга, и различимы при использовании масштаба в 3 мм. ЭЭГ, зарегистрированная со скальпа, имеет пространственное разрешение, равное нескольким сантиметрам. Обратите внимание на 10-кратное различие масштабов амплитуды потенциалов, зарегистрированных интракортикально и с поверхности коры.
В настоящее время известно, что работа мозга определяется совместным функционированием нейронных ансамблей, а не активностью одиночных нейронов. Но 40 лет назад ученые придерживались противоположной точки зрения, предполагая, что только исследование поведения отдельных нейронов сможет дать ключ к пониманию функций всего мозга. В те дни возник новый метод — регистрация активности одиночного нейрона (рис. 1). Большинство нейрофизиологов покинуло область классической ЭЭГ (единственного доступного в те дни электрофизиологического метода) и приступило к исследованию активности одиночных нейронов.
В некоторой степени ожидания были оправданы. Действительно, открытие свойств простых и сложных клеток зрительной коры Хюбелем и Визелем (награжденными Нобелевской премией в 1981 г.) казалось, подтвердило предположение о том, что изучение поведения одиночных нейронов вносит новый вклад в понимание процессов обработки информации в мозге животных.
Регистрация активности нейронов
В 1970-х, изучая реакции одиночных нейронов и нейронных популяций мозга человека с помощью крошечных электродов, имплантированных в различные области мозга неврологических и психиатрических пациентов, мы случайно обнаружили сходство нейрональных реакций в некоторых отделах мозга (рис. 2). Оно выражалось в подобии так называемых профилей нейрональных реакций — усредненных изображений ответов всех нейронов, зарегистрированных в определенной анатомической структуре. Сравнение профилей реакций с локальными потенциалами, зарегистрированными теми же имплантированными электродами, показало, что коллективное поведение нейронов находит довольно точное отражение в локальных потенциалах (рис. 3).
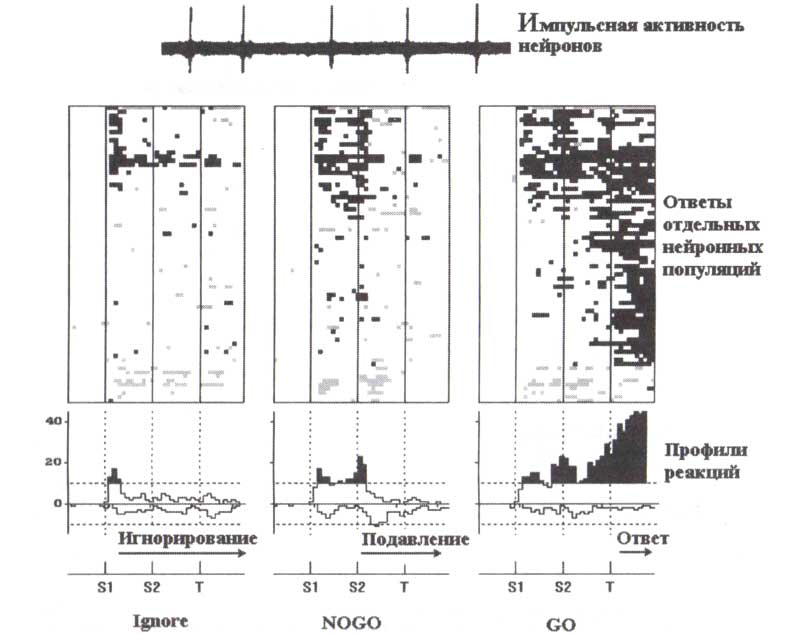
Активность нейронной популяции (вверху) при выполнении двухстимульного GO/NOGO-теста оценивалась с помощью электродрр, имплантированных в базальные ганглии и таламус пациентов. Каждая горизонтальная линия (посередине) соответствует отдельной нейронной популяции. Черные (серые) клетки: значимое (р < 0,01) увеличение (уменьшение) частоты разрядов по отношению к фоновой частоте разрядов. Бинарные постстимульные гистограммы были усреднены для получения профилей реакций (внизу). Вертикальные оси: число нейронных популяций, значимо активированных (выше) или инактивированных (ниже); пунктирные горизонтальные линии: уровни значимости р < 0,01. Горизонтальные оси: время в 100 мс bins. Слева направо представлены типы проб: игнорируемые, NOGO- и GO-пробы. S1, S2, Т — предъявления первого, второго и третьего стимулов в пробах. Заимствовано из Kropotov et al. (1997).
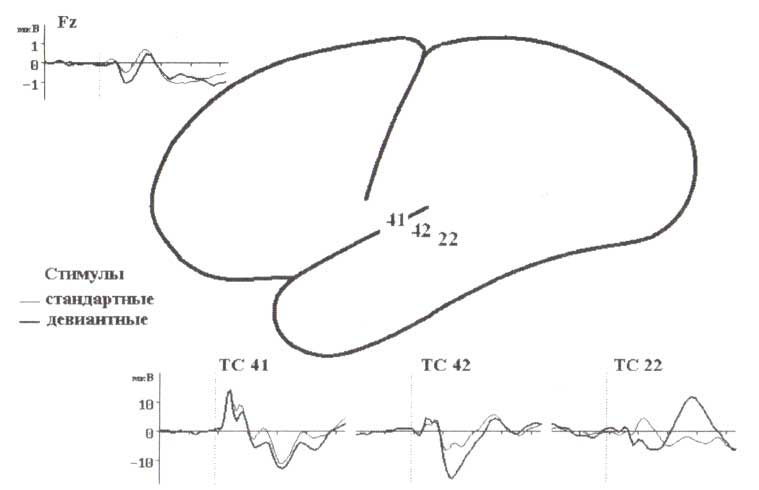
В тесте стандартные акустические стимулы (тон частотой 1000 Гц и длительностью 100 мс) предъявлялись последовательно с интервалами в 800 мс и в случайном порядке смешивались с редкими (вероятность — 15 %) девиантными (тон частотой 1300 Гц) стимулами. Испытуемые читали книгу. Обратите внимание на 10-кратное различие амплитуды сигналов, зарегистрированных со скальпа и интракортикально. Заметьте также, что паттерны ответов различны в трех разных слуховых зонах: поле Бродмана 41 — первичная слуховая кора, поле Бродмана 42 — вторичная слуховая кора, поле Бродмана 22 — ассоциативная слуховая зона.
Локальные потенциалы
Основная статья: Локальные потенциалы
Помня об этих результатах, в начале 90-х мы стали использовать регистрацию локальных потенциалов для анализа этапов обработки информации в мозге человека. Работа проводилась совместно с Р. Наатаненом и его коллегами из Отдела исследований головного мозга (Cerebral Brain Research Unit (CBRU)) Университета г. Хельсинки. Во время этих исследований ученые обнаружили, что определенные области слуховой коры генерируют различные вызванные потенциалы, что дало возможность изучать детали обработки информации в слуховой коре человека. Действительно, первичное слуховое поле 41 -кодировало в своих вызванных локальных потенциалах частоты формант акустических стимулов в диапазоне ранних (до 50 мс) временных интервалов, но не обнаруживало никаких эффектов -памяти, т.е. не проявляло привыкания к повторяющимся слуховым стимулам. В противоположность этому для ответов вторичного слухового поля 42 было характерно существенное привыкание, что доказывало наличие выраженного эффекта-памяти в этой области. Самые впечатляющие ответы генерировались в слуховом ассоциативном поле 22, где возникал дополнительный потенциал в случае, если стимул не соответствовал ряду предшествовавших монотонно повторяющихся стимулов. Комбинация этих трех типов локальных потенциалов, по-видимому, находит отражение в регистрируемом с поверхности головы негативном колебании, названном Mismatch Negativity.
Взаимосвязь локальных потенциалов и скальповой ЭЭГ
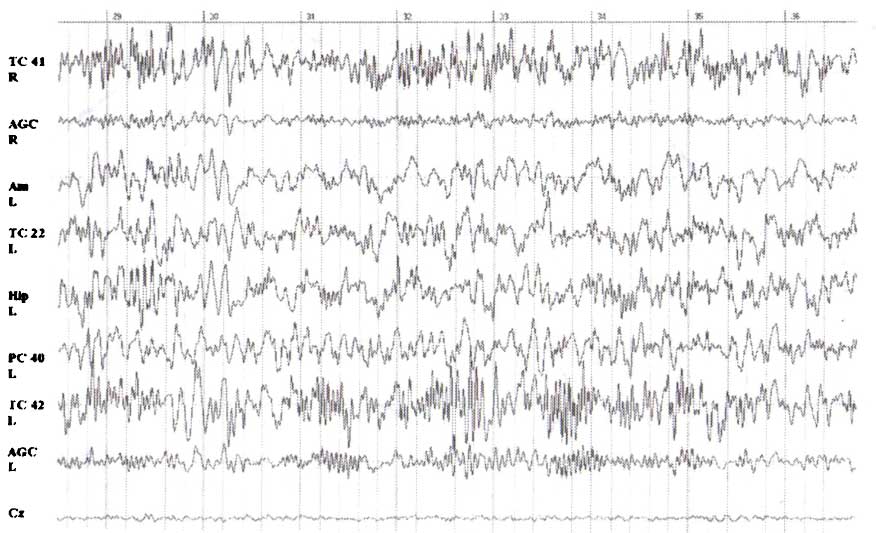
Спонтанные локальные потенциалы, зарегистрированные с помощью интракортикальных электродов в различных областях мозга (ПБ 41, 46, 42, 22, гиппокамп, миндалина, передняя цингулярная извилина) по сравнению с ЭЭГ, зарегистрированной со скальпа в точке Cz. Обратите внимание на то, что в различных зонах генерируются различные паттерны локальных потенциалов. В среднем они в 10 раз превосходят по величине скальповую ЭЭГ, довольно независимы и отличаются по доминирующей частоте.
Результаты исследований представлены на рис. 3 и отражают два явления: I) усредненные по всем пробам локальные потенциалы дают детальную информацию о временном и пространственном распределении локальных операций в нейронных сетях коры; 2) локальные потенциалы находят отражение в когнитивных вызванных потенциалах, регистрируемых с поверхности головы, со значительно меньшими амплитудными значениями. Сходные выводы могут быть сделаны относительно взаимосвязи спонтанной ЭЭГ и интракортикальных локальных потенциалов. В нашей лаборатории при изучении спонтанных локальных потенциалов, зарегистрированных с имплантированных электродов, мы столкнулись с тем фактом, что в различных полях Бродмана выяляются довольно отличающиеся паттерны колебаний. Пример такой записи у пациента с вживленными электродами представлен на рис. 4.
Возрождение ЭЭГ в наши дни
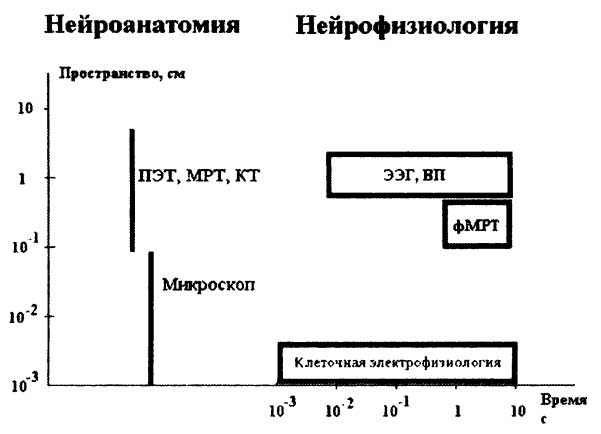
Пространственное (ось Y) и временное (ось X) разрешения изображены в виде прямоугольников. Обратите внимание на отсутствие временного разрешения в техниках нейроанатомии. Аббревиатуры: компьютерная томография (КТ), позитронная эмиссионная томография, магнитно-резонансная томография. Нейроанатомия связана с различными пространственными шкалами. Пространственное разрешение микроскопа меньше размера клетки (около 30 микрон — 3 х 10-3 см). ПЭТ и МРТ дают гораздо меньшее пространственное разрешение. Нейрофизиологические параметры включают электрические и электромагнитные свойства нейронов и нейронных сетей и служат предметом изучения клеточной электрофизиологии, скальповой ЭЭГ и когнитивных ВП.
Несмотря на то что метод ЭЭГ возник почти 80 лет назад, самые быстрые темпы его развития наблюдаются только в последнее время. В наши дни мы видим возрождение ЭЭГ Существует по меньшей мере четыре причины этого явления. 1 кривая связана с недавним появлением новых методов анализа ЭЭГ, таких как техника пространственной фильтрации при коррекции артефактов, анализ независимых компонент когнитивных ВП, вэйвлет-анализ, электромагнитная томография и некоторые другие методы. Вторая причина заключается в относительной дешевизне современных электроэнцефалографов. Действительно, в наши дни приборы для регистрации ЭЭГ стоят от нескольких тысяч до нескольких десятков тысяч долларов США, что недорого по сравнению с многомиллионной стоимостью оборудования для МРТ и ПЭТ. Третья причина — значительный рост наших познаний о механизмах генерации волн спонтанной ЭЭГ и функционального значения компонентов когнитивных ВП. И, наконец, четвертая — высокое временное разрешение сигналов ЭЭГ и когнитивных ВП, что принципиально не может быть достигнуто другими техниками нейрокартирования. Методы ЭЭГ и когнитивных ВП обеспечивают временное разрешение сигналов в несколько миллисекунд, в то время как ПЭТ и МРТ ограничены разрешением в несколько секунд.