Функциональный смысл детекции изменений
Окружающий мир отражается в постоянной активности нейронов сенсорных систем мозга. Некоторые части мира остаются неизменными в течение достаточно длительных периодов времени, на которые мы чаще всего не обращаем внимания. Но когда нас вдруг кто-нибудь трогает за рукав, или нарушается регулярность работы автомобильного двигателя, или на глаза попадается гриб в лесу, эти изменения могут быть сознательно замечены. По-видимому, мозг постоянно сравнивает сенсорную картину мира с его моделью, сформированной на основе предварительной стимуляции. Похожим образом мозг контролирует реализацию действий и, если они не соответствуют запланированным, корректирует поведение. Мы будем называть эти базовые операции операциями сравнения. Исторически в сенсорной модальности операция сравнения называется детекцией.
Два типа операций сравнения соответствуют двум разновидностям моделей окружающего мира: сенсорной и моторной. Модели формируются при участии сенсорной и исполнительной систем мозга. Несмотря на сильную взаимосвязь между ними, каждая из этих систем представляет собой отдельную нейронную сеть с определенным механизмом переработки информации, служащую для реализации определенной функции. Соответственно, в поведении человека детекция изменений является важной для достижения двух основных целей.
Во-первых, изменения в сенсорной картине мира привлекают внимание как сознательно (селективное внимание), так и бессознательно (автоматически). Например, неожиданное изменение в шуме работающего двигателя может быть замечено водителем и когда внимание сконцентрировано на дороге, и при намеренном прослушивании. В этой ситуации сенсорная модель формируется на базе сенсорной системы, а текущий стимул, не соответствующий данной модели, вызывает активацию соответствующей части сенсорной системы. Важную роль в формировании модели играет и предыдущий сенсорный опыт. Таким образом, цель данного типа детекции изменений обусловлена функцией сенсорной системы — выбор источника изменения сенсорной информации для ее более детальной обработки.
Во-вторых, факт детекции изменения может означать, что реализованное действие не является корректным, а значит, должно быть изменено или подавляться в дальнейшем. Так, человек, собирающийся переходить улицу на зеленый свет, останавливается при резкой смене сигнала светофора на красный. В этой ситуации модель действия — переходить дорогу — формируется на уровне исполнительной системы. Изменение текущей поведенческой ситуации означает; что подготавливаемое действие является несоответствующим и должно быть подавлено. Таким образом, основной целью данного типа детекции изменений является подавление исполнительной системы для обеспечения отмены реализации планируемого действия.
Механизм детекции изменений (модель)
Наиболее изученными являются механизмы мониторинга изменений окружающей среды в слуховой модальности, с рассмотрения которой мы и начнем. Лидирующие позиции в исследовании этого вопроса занимают исследования Центра когнитивных исследований мозга Университета Хельсинки (Финляндия), созданного Ристо Наатаненом в 1980-х годах. Наатанен был первым исследователем, предположившим существование двух механизмов детекции изменений для слуховой модальности.
Первый механизм отражается изменениями ранних компонент ВП и ассоциируется с обработкой информации на уровне первичной и, скорее всего, вторичной слуховой коры (рис. 1). Работу первого механизма Ристо Наатанен связывает с N1 компонентой ВП. Предполагается, что в составе слуховой коры существует две разновидности нейронов — стабильные и рефрактерные. Стабильные нейроны не меняют свои ответы при повторных появлениях стимула. Активность рефрактерных нейронов снижается по мере того, как несколько раз предъявляется один и тот же стимул. Рефрактерность означает феномен снижения частоты ответов при повторяющейся стимуляции. Хотя синаптическая депрессия, как кажется, является основным клеточным механизмом затухания нейронных ответов на повторяющиеся стимулы, сложные взаимодействия между нейронами, по-видимому, также играют некоторую роль (пока еще неопределенную) в реализации этого феномена.
Кроме существования двух типов нейронов, предполагается наличие и второго механизма, называемого детекцией изменений. Гипотетические нейроны, ответственные за обеспечение работы механизма, реагируют только на изменения стимулов. Если принять во внимание нелинейный характер активности нейронов, то легко можно представить возможное устройство нейронной сети для детекции изменений (рис. 1 справа). Исходя из предложенной модели, ассоциативная кора (кортикальная область 3 на рис. 1) получает два входа: 1) из первичной сенсорной коры, ответственной за обработку физических свойств стимула и не изменяющей своей активности при повторении стимула; 2) из вторичной, активность которой снижается при повторении. Нейроны ассоциативной области посредством латерального торможения осуществляют сравнение информации, поступающей с этих двух входов. В результате некоторые нейроны этой области активируются только при появлении нового стимула.
Аудиостимулы характеризуются не только физическими свойствами (частотой и интенсивностью), но и более интегративными свойствами, например, таким как регулярность. В частности, в слуховой модальности регулярность представлена во множестве звуковых сигналов нашей повседневной жизни: шуме работающего автомобильного двигателя, компьютерного вентилятора или монотонной речи лектора… Как известно из нейрофизиологии, разные аспекты стимула обрабатываются нейронными элементами, локализованными в различных областях сенсорных систем. Регулярность как отдельное свойство стимула, по всей видимости, кодируется обособленной областью слуховой коры. Более того, весьма вероятно, что это свойство оценивается с помощью специального мозгового механизма. В действительности в отличие от физических характеристик регулярность как интегративное свойство формируется на основе достаточно длительного отрезка времени (десятков секунд). Однако механизм этой интеграции неизвестен. Несмотря на это, ЭЭГ-корреляты реакции мозга на изменение регулярности за последние 30 лет хорошо исследованы. Для слуховой модальности этим коррелятом является негативность рассогласования (HP).
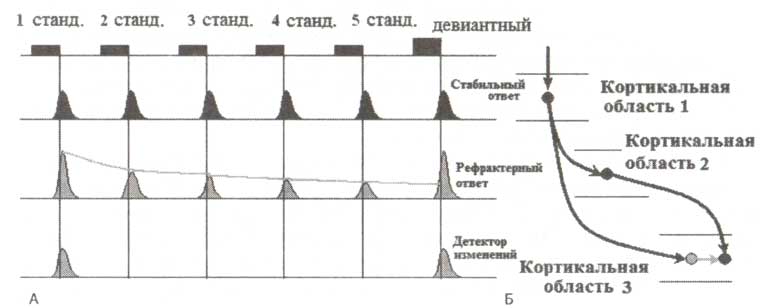
А. Нейрональные ответы. Б. Связи между нейронами трех предполагаемых областей коры. Черным цветом обозначены возбуждающие нейроны и связи, а серым — тормозные нейроны и связи. В соответствии с представленной моделью предполагается, что операции сравнения осуществляются сетью нейронов, состоящей из трех кортикальных областей. Первая из этих областей получает входы непосредственно от таламуса, и ее активность является наиболее устойчивой, т.е. стабильна при повторении стимулов. Далее проекции от этой области следуют на возбудительные нейроны второй области, активность которой более пластична, и тормозные нейроны третьей области, где осуществляется детекция изменений (т.е. область детекторов изменений). В свою очередь, нейроны третьей области, которые находятся под воздействием локальных тормозных нейронов, получают входы от возбудительных нейронов второй области. Таким образом, происходит сравнение, т.е. вычисление разности двух входов — пластичного и более ригидного. Если предъявляемый стимул является новым, то возбуждение превышает торможение, что приводит к разрядке нейронов. В обратном случае, когда стимулы повторяются, наблюдается преобладание торможения над возбуждением, и нейроны не реагируют на такие стимулы. Обозначения: станд. — стандартный стимул.
HP в oddball-парадигме
Негативность рассогласования — электрический ответ на различаемое изменение регулярности слуховой стимуляции. Обычно этот ответ регистрируется на ВП с латентностью пика 150—200 мс с момента начала изменений, даже при отвлечении внимания. Феномен HP впервые был продемонстрирован для слуховой модальности, а его наличие в зрительной модальности является спорным вопросом. Причина этого заключается в природе операции сравнения, которая основана на сравнении со следом памяти. След в краткосрочной сенсорной памяти формируется в результате процесса реверберации и синаптической депрессии (или потенциации). Известно, что эхоическая память достаточно длительна — до 10 с, тогда как иконическая крайне коротка — до нескольких сотен миллисекунд. Таким образом, регулярность может быть сформирована в рамках традиционной исследовательской oddball парадигмы в слуховой, но не зрительной модальности. Следует подчеркнуть, что HP не регистрируется, если межстимульный интервал длится больше чем 10 с, что соответствует представлениям о временных характеристиках краткосрочной сенсорной памяти.
Изучение HP в слуховой модальности традиционно проводится с использованием так называемой oddball-парадигмы. В упрощенном варианте тестовое задание выглядит следующим образом: в ряду повторяющихся стандартных стимулов (например, тон 1000 Гц длительностью 100 мс) редко предъявляются девиантные (например, тон 1100 Гц длительностью 100 мс). Более детальное описание приведено в главе «Методы» части 2.
Феномен негативности рассогласования был открыт в 1978 году в классической работе Наатанена, Джилларда (Gaillard) и Мантисало (Mantysalo). В соответствии с общепринятой интерпретацией, HP генерируется в результате детекции рассогласования между девиантным стимулом и следом памяти, сформированным повторно предъявляемыми стандартными слуховыми стимулами. Усредненные ВП, зарегистрированные в рамках oddball-парадигмы при исследовании здоровых испытуемых в возрасте от 15 до 89 лет, представлены на рис. 2 (слева). По сравнению со стандартными стимулами предъявление девиантных вызывало увеличение амплитуды негативного отклонения потенциала в области лобно-центральных отведений.
С использованием метода независимых компонент было произведено разложение полученных ВП и выделены три независимых компоненты (рис. 2 справа). Первая компонента генерировалась в области первичной слуховой коры с латентностью пика 116 мс. Фактически эта компонента была одинакова и для стандартного, и для девиантного стимулов. Вторая компонента, генерировавшаяся в ассоциативных областях слуховой коры с латентностью пика 132 мс, характеризовалась вдвое большей амплитудой для девиантного стимула по сравнению со стандартным. Наконец, третья компонента была обнаружена в области передней поясной извилины. Латентность его пика составляла 236 мс. Компонента при девиантном стимуле (относительно стандартного) характеризовалась дополнительной позитивностью.
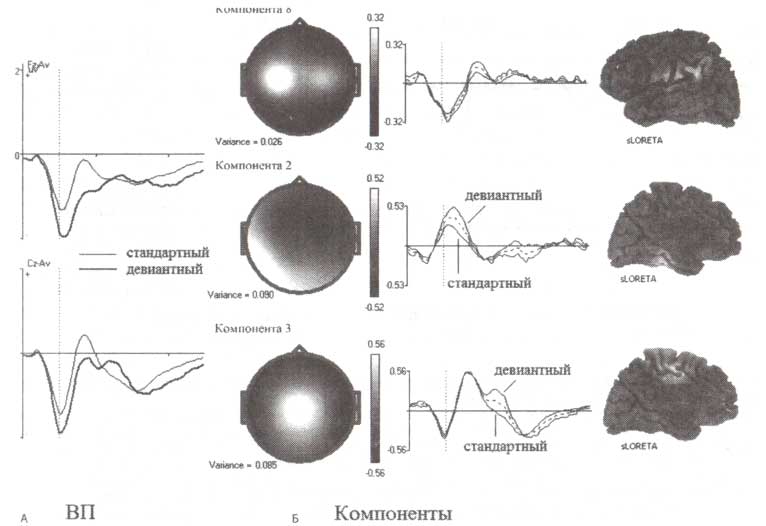
А. Усредненные по группе здоровых добровольцев ВП. Б. Топография, временная динамика и локализация генераторов трех независимых компонент, составляющих полученные ВП, вычисленная с помощью метода томографии низкого разрешения (sLORETA).
Внутримозговые корреляты HP
В исследовании, проведенном в нашей лаборатории совместно с финским коллективом, возглавляемым Р. Наатаненом (Kropotov et al., 2000), было показано существование по крайней мере трех отдельных мозговых механизмов генерации HP. На рис. 3 представлены локальные потенциалы, зарегистрированные с электродов, имплантированных в слуховую кору в области 41-, 42- и 22-го ПБ. Как видно из рис. 3, стабильная активность демонстрируется только для первичной слуховой коры (ПБ 41) и не изменяется при повторно предъявляющихся стимулах. Следовательно, можно считать, что нейроны этой области обеспечивают кодирование физических характеристик стимулов (например, частоту сигнала). Вторичная слуховая кора (ПБ 42) снижает активность при повторении одинаковых стимулов. Вполне вероятно, что определенный вклад в наблюдаемый эффект может вносить синаптическая депрессия. По всей видимости, операция сравнения происходит на уровне ассоциативной области слуховой коры (ПБ 22): если стимул не соответствует следу памяти, сформированному предварительной однотипной стимуляцией, то генерируется сильно выраженная негативная компонента, за которой следует позитивная. Следует оговориться, что слуховая кора не является единственной областью, влияющей на генерацию HP. Так, кроме височной области, были найдены несколько источников генерации в префронтальной коре. Генераторы HP, локализуемые в лобных долях, активируются после височных, спустя небольшой промежуток времени и играми важную роль в процессах инициации бессознательного переключения внимания на изменение звука, детектируемого в височном коре. Предполагается, что процесс детекции изменения происходит именно в слуховой коре (отражается субкомпонентом HP в нисочной области). Это является триггером активации фронтальных областей коры (генерируется фронтальный субкомпонент HP и Р3а компонент, следующий за ним), которые, вероятно, обеспечивают переключение внимания и сознательное восприятие изменения стимула.
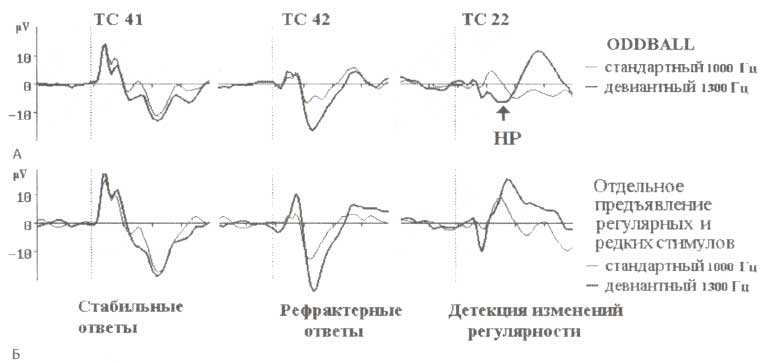
Внутримозговые локальные полевые потенциалы регистрировались у пациентов, страдающих эпилепсией, которым по клиническим показаниям, с целью диагностики и лечения, в различные области височной доли имплантировались долгосрочные электроды (области Бродмана 41,42 и 22). В одном исследовании активность нейронов регистрировалась в момент чтения книги пациентом, при параллельном предъявлении последовательностей тонов. А. Тоны предъявлялись в рамках oddball-парадигмы: стандартные тоны 1000 Гц были перемешаны с девиантными 1300 Гц. В другом исследовании (Б) предъявлялись только стандартные и девиантные стимулы с соответствующими длинными и короткими межстимульными интервалами (Kropotov etal., 2000). Обозначения: HP — негативность рассогласования; ТС — область слуховой коры.
Детекция изменений в задаче по дискриминации двух стимулов
Негативность рассогласования представляется как частный случай более общей операции, ассоциирующийся с детекцией изменения регулярности звуковой стимуляции. Однако регулярность как свойство сенсорной стимуляции наиболее выражена именно в слуховой модальности.
В общем смысле операция сравнения осуществляется с любой характеристикой сенсорного стимула и соответствующим следом памяти. Мы называем эту операцию детекцией изменения. Если задачей исследования не является изучение процессов, связанных с неожиданными изменениями регулярности стимулов, то нет необходимости использовать задание со стимулами, повторяющимися много раз. Следовательно, детекция изменений может быть экспериментально исследована не только в рамках oddball-парадигмы. Так, нами было разработано тестовое задание по дискриминации двух стимулов для изучения процессов детекции изменений свойств зрительной стимуляции. В этом тестовом задании стимулы предъявлялись парами, в которых второй стимул мог в некотором смысле совпадать или не совпадать с первым, соответственно задачей испытуемого было по-разному реагировать на пары проб этих двух категорий.
В исследовании обезьян (Romo et al., 2000) изучались процессы различения поверхностей при регистрации импульсной активности нейронов (ИАН), расположенных в соматосенсорных областях S1 и S2, вентральной префронтальной и медиальной премоторной областях коры. Для этого на кончике одного из пальцев размещался механический стимулятор, производивший короткие колебания. Каждая проба состояла из стимуляций с разными частотами f1 и f2, разделенными коротким интервалом времени. Обезьяна должна была нажимать определенные кнопки в зависимости от того, была ли частота стимуляции f2 выше или ниже, чем f1. Последовательность психологических операций представлялась следующим образом:
- кодирование частоты первой вибрации f1;
- удержание следа в сенсорной памяти;
- кодирование частоты второй вибрации f2;
- сравнение результата кодирования со следом памяти f1;
- передача результата операции сравнения моторным нейронам.
Полученные в этом исследовании результаты свидетельствовали о том, что операция сравнения обеспечивалась нейронами вторичной (а не первичной) соматосенсорной коры. То есть «вычислялась» разница между двумя типами вибраций, которая отражалась в частоте разрядов этих нейронов в ответ на второй из пары стимулов пробы. Время, необходимое для реализации этой операции, составляло 200 мс. Доказательств участия нейронов первичной сенсомоторной коры (S1) в обеспечении операции сравнения выявить не удалось, а профиль их ответов выглядел как функция зависимости от частоты второго стимула (f2).
Модальная специфичность
Начиная с 1990-х годов операции сравнения являются предметом исследований нашей лаборатории. Нами была разработана так называемая двухстимульная GO/NOGO-парадигма. Исследования проводились как в слуховой, так и в зрительной модальности. В качестве слуховых стимулов использовались высокие (В) и низкие тона (Н) в следующих сочетаниях: ВВ, ВН, НВ, НН. В качестве зрительных стимулов использовались цифры 6 и 9 в сочетаниях: 99, 96, 69, 66. Задачей испытуемого являлось выделять пары стимулов и нажимать кнопку, если второй стимул в паре (высокий тон или цифра 9) соответствовал первому (высокий тон или 9 соответственно). Мы полагали, что после инструкции и тренировочной сессии у испытуемого формируется поведенческая модель соответствия пары стимулов выбранной категории (например, два изображения цифры 9) с определенным действием (нажатие кнопки). Считалось, что модель состоит из двух частей: 1) сенсорная модель — шаблон категории релевантных стимулов;2) моторная модель — шаблон действия (нажатие кнопки), которое должно быть исполнено как можно быстрее и точнее.
Выполнение тестового задания состояло из ряда психологических операций. Предъявление первого стимула индуцировало:
- кодирование релевантных характеристик стимула;
- сравнение характеристик с сенсорным шаблоном;
- в случае совпадения — обновление сенсорной подготовительной установки (preparatory set) и инициация моторной подготовительной установки;
- в случае рассогласования — подавление формирования моторной подготовительной установки.
Психологические операции, вызываемые предъявлением второго стимула пары (GO или NOGO):
- кодирование релевантных свойств стимула;
- сравнение эти свойств с характеристиками предыдущего (первого) стимула;
- в случае совпадения — инициация выбора действия и установочной реакции;
- при рассогласовании — подавление процессов автоматического выбора действия и обновление установочной реакции;
- сравнение результатов NOGO-пробы (когда действие не реализуется) с шаблоном действия (когда следует нажимать кнопку) и коррекция дальнейшего поведения.
Для упрощения мы рассмотрим ВП только при предъявлении вторых стимулов проб. Анализироваться будут только первые этапы обработки сенсорной информации. Ранними отличиями между GO и NOGO являются: 1) модальная специфичность; 2) разный компонентный состав (пространственно перекрывающиеся, но разделенные по времени), отражающий соответственно физические и смысловые изменения стимулов; 3) отличие от компонент, связанных с моторной реакцией.
На рис. 4 показано, что ранние компоненты разностных ВП для GO- и NOGO-состояний модально специфичны. Для зрительных стимулов максимальная позитивность регистрировалась в области Т5-отведения, тогда как в слуховой модальности максимальные значения компоненты наблюдались в F7. Следует отметить и большую скорость обработки слуховой информации: пиковая латентность разностного ВП в слуховой модальности — 200 мс, а в зрительной — 240 мс.
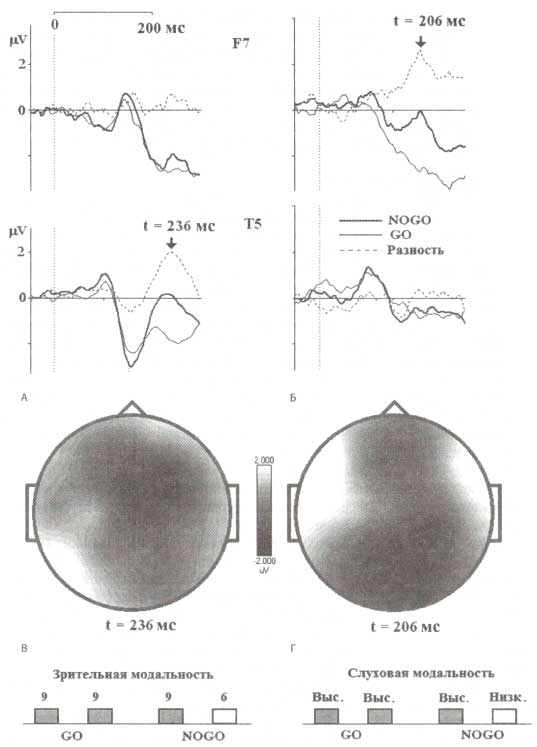
В верхней части расположены графики скальповых ВП для проб GO (тонкая линия) и NOGO (толстая линия), а также разностных ВП NOGO-GO (пунктирная линия), которые были зарегистрированы в F7- и Т5-отведениях. Слева представлены ВП в зрительной модальности, а справа — в слуховой. В нижней части рисунка представлены топограммы разностных ВП. Обозначения: выс., низк. — тоны высокой и низкой частоты.
Детекция физических и смысловых изменений
Рис. 5 иллюстрирует тот факт, что ранние компоненты разностных ВП для GO- и NOGO-состояний действительно слагаются из двух составляющих. Исследования проводились на больных эпилепсией, которым были вживлены электроды в целях диагностики и терапии. Исследование осуществлялось в двухстимульной парадигме в разные дни: в один из дней требовалось нажимать кнопку только в пробах с парой слуховых стимулов ВВ (GO-условие), в другой при предъявлении ВН-пары (GO-условие). В результате NOGO-условие в обоих случаях характеризовалось позитивной компонентой, регистрируемой с ЛП около 200 мс. В то же время ранняя негативная компонента с ЛП 100 мс генерировалась в ответ на изменение физических свойств ожидаемого стимула. Следует также отметить, что ранняя негативная компонента регистрировалась и в ситуации, когда первый стимул предъявляемой пары не соответствовал ожидаемому (т.е. Н-стимул на первом месте в паре вместо ожидаемого В-стимула).

Запись, сделанная с имплантированных электродов, расположенных в средней височной доле (вверху) у пациента, страдающего эпилепсией. Электрическая активность, регистрировавшаяся в течение двух разных дней, представлена в верхней и средней области рисунка. Пациент выполнял двухстимульный слуховой тест, в котором с равной вероятностью в случайном порядке предъявлялись тоны высокой (Н) и низкой частоты (L). В течение первого исследовательского дня задачей пациента являлось нажимать кнопку в случае предъявления пары стимулов высокой частоты (НН). На второй день исследования нужно было нажимать кнопку при парном предъявлении стимулов высокого и низкого тонов (HL).
Детекция изменений и подавление моторной реакции
Для демонстрации различий в топографии компонент, связанных с детекцией изменений и подавления моторного ответа, мы рассмотрим результаты анализа зрительных ВП в GO/NOGO-задании, выполненном с помощью метода независимых компонент (рис. 6). Как видно из рис. 6, первая из наиболее выраженных компонент генерируется в левой височной области и, вероятно, отражает детекцию как физических, так и смысловых изменений стимулов. Вторая компонента генерируется в области левой премоторной коры и, по всей видимости, характеризует процессы подавления выполнения подготовленного действия.
Сравнительно недавно группой ученых из Университета Хоккайдо в Японии (Kimura et al., 2006) в исследовании, проводимом в двухстимульной парадигме (SI—S2) в зрительной модальности, были зарегистрированы ВП, связанные с детекцией изменений. Как и в наших работах, в этом исследовании было показано, что изменение стимулов сопровождалось генерацией позитивного компонента с ЛП 100—200 мс в задних областях мозга. Варьирование различных характеристик зрительных стимулов (детекция изменения цвета и направления движения) характеризовалось специфическими топографическими и временными паттернами распределения компонент.
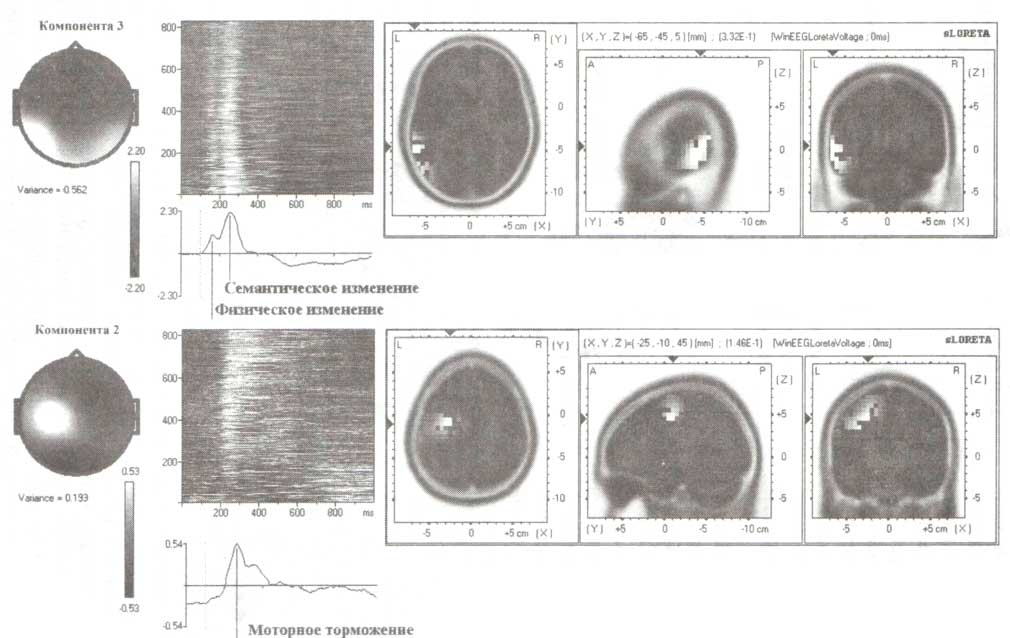
Представлены результаты анализа разностного ВП (GO-NOGO), произведенного методом независимых компонент. Изображены данные для более чем 800 здоровых испытуемых возрастом от 7 до 89 лет. Слева направо: 1) топография компонент и значения их вариабельности (под топограммами); 2) вертикально уложенные тонкие горизонтальные окрашенные полосы (цвет обозначает амплитуду), каждая из которых представляет изменение во времени амплитуды соответствующей компоненты ВП отдельного испытуемого: представлены данные приблизительно 800 здоровых испытуемых возрастом от 7 (расположены внизу) до 89 лет (расположены вверху); 3) временная шкала компонент со стрелками; 4) локализация генераторов компонент, вычисленная с помощью метода томографии низкого разрешения (sLORETA). Шкалы представлены вблизи соответствующих изображений.