Ионные токи, потенциал покоя и потенциал действия. Распространение потенциала действия по аксону.
Биопотенциалы – это электрическое напряжение, которое генерируется физиологическими процессами, происходящими в организме. Биопотенциалы создаются электрохимической активностью клетки, называемой возбудимой клеткой. Когда возбудимая клетка подвергается внешним стимулам, она генерирует потенциал действия, который является основным источником биопотенциалов в организме.
Биопотенциалы головного мозга
Биоэлектрическая активность мозга представляет собой процесс, обусловленный активностью огромного числа генераторов-нейронов, и, в соответствии с этим, создаваемое ими поле представляется весьма неоднородным по всему пространству мозга и меняющимся во времени. В связи с этим между двумя точками мозга, а также между мозгом и удаленными от него тканями организма возникают переменные разности потенциалов, регистрация которых и составляет задачу электроэнцефалографии.
Биопотенциал, отражающий электрическую активность мозга, называется электроэнцефалограммой. Необходима достаточная продолжительность и длина небольших по величине (микровольты) электрических потоков биоэлектрической активности мозга, для того чтобы сигнал можно было усилить и сделать доступным для интерпретации.
ЭЭГ, а также записи, сделанные с помощью электродов, имплантированных в мозг животных, показывают, что мозг живого животного постоянно активен, даже во время сна. Каждая часть мозга демонстрирует смесь ритмической и неритмической активности, которая может варьироваться в зависимости от состояния и поведения. Идентификация источника(ов) генератора и распространения электрического поля (полей) служит основой для распознавания электрографических паттернов, которые определяют проявления биоэлектрической активности головного мозга как нормальные или патологические. В большинстве случаев рутинная ЭЭГ регистрируется с поверхности головы и отражает суммарную электрическую активность, генерируемую множеством нейронов.
Происхождение биопотенциалов
Возбудимые типы клеток, которые способствуют генерации биопотенциалов:
- Афферентные (рецепторные или сенсорные) нейроны, которые передают сигналы от тканей и органов к центральной нервной системе
- Эфферентные (моторные) нейроны, которые передают сигналы от центральной нервной системы к эффекторным клеткам
- Эффекторные клетки, которые включают мышечные клетки и нейроэндокринные клетки, которые стимулируют физическое воздействие на основе полученного сигнала
- Интернейроны, которые полностью существуют в центральной нервной системе, включая мозг. Интернейроны передают сигналы между афферентными нейронами и эфферентными нейронами.
Механизм потенциала действия
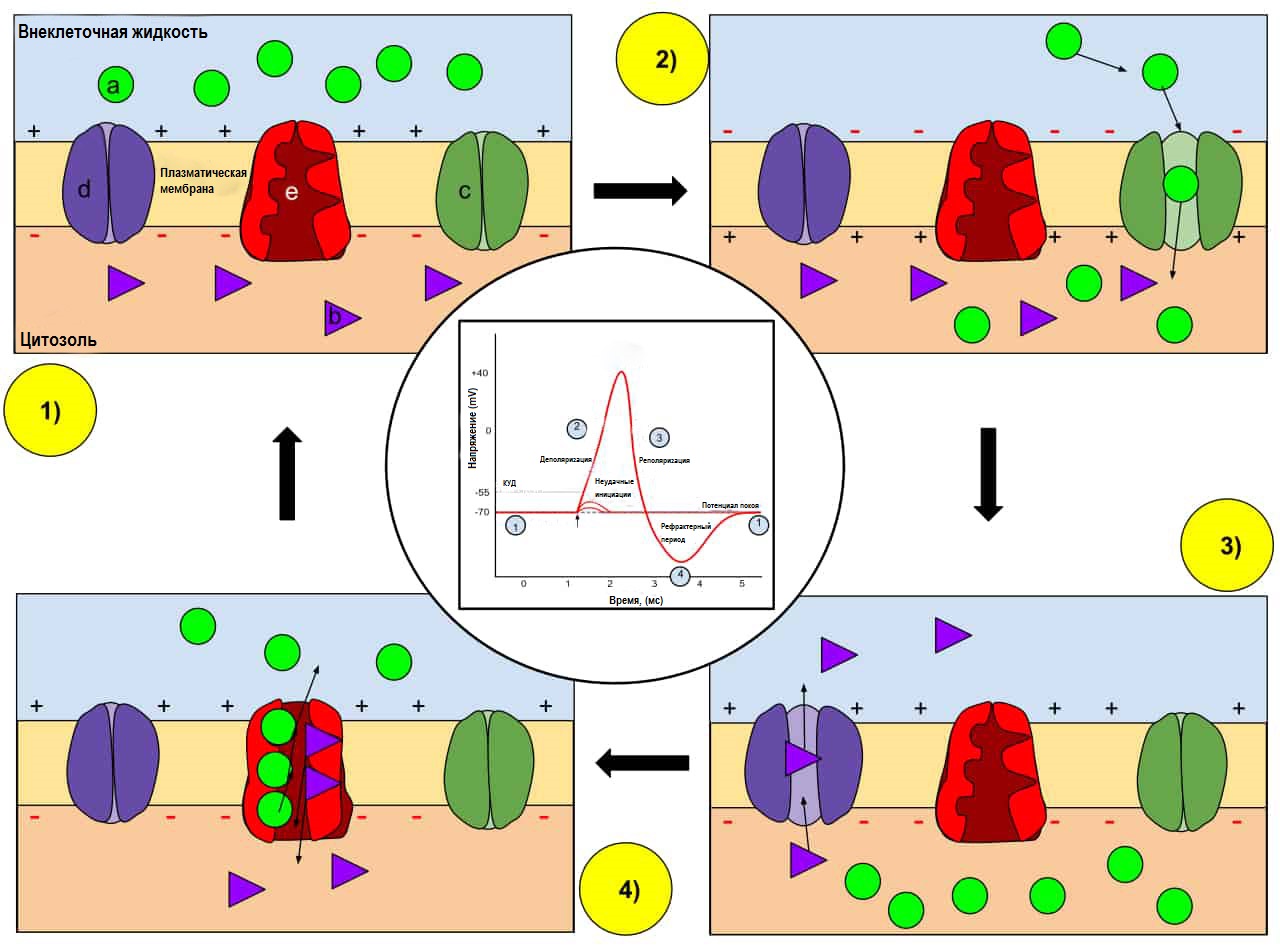
а) Ион натрия (Na +); b) Ион калия (К +); c) Натриевый канал; d) Калиевый канал; e) Натриево-калиевый насос. В момент потенциала действия проницаемость мембраны нейрона изменяется. В состоянии покоя (1) ионы натрия и калия имеют ограниченную способность проходить через мембрану, а нейрон имеет чистый отрицательный заряд на внутренней стороне мембраны. После запуска потенциала действия деполяризация (2) нейрона активирует натриевые каналы, позволяя ионам натрия проходить через клеточную мембрану в клетку, что приводит к изменению отрицательного заряда внутренней стороны мембраны на положительный относительно внеклеточной жидкости. После достижения пика, нейрон начинается фаза реполяризации (3), где натриевые каналы закрываются, а калиевые каналы открываются, позволяя ионам калия проникать через мембрану во внеклеточную жидкость, возвращая потенциал мембраны к отрицательному значению. Наконец, существует рефрактерный период (4), в течение которого зависимые от напряжения ионные каналы инактивируются, в то время как ионы Na + и K + возвращаются к своим распределениям в момент состояния покоя (1), и нейрон готов к генерации следующего потенциала действия.
Нейроны порождают электрические импульсы, что связано с так называемым потенциалом действия . Это явление связано с током положительно заряженных ионов через мембрану нейрона. Ионный ток обусловлен наличием в клеточной мембране особых типов потенциалзависимых ионных каналов (натриевые и калиевые). Потенциалзависимые натриевые каналы относятся к типам каналов, способных создавать положительную обратную связь, необходимую для создания потенциала действия, т.е. мембранный потенциал управляет состоянием ионных каналов, как и состояние ионных каналов управляет мембранным потенциалом.
После воздействия стимула происходит смещение мембранного потенциала. При этом открываются натриевые ионные каналы и положительно заряженные ионы натрия по электрохимическому градиенту проникают в нейрон и частично нейтрализуют отрицательный на внутренней стороне мембраны – начинается деполяризация. После этого активируются потенциалзависимые натриевые каналы, поток ионов натрия усиливается, что вызывает нарастание скорости деполяризации. После достижения пика натриевые каналы начинают постепенно закрываться, а калиевые ионные каналы открываются, позволяя выводить ионы калия из клетки, возвращая мембранный потенциал к уровню покоя – фаза реполяризции. Цикл деполяризации и реполяризации является чрезвычайно быстрым, занимая всего около 2 миллисекунд (0,002 секунды) и, таким образом, позволяет нейронам запускать потенциалы действия при быстрых спайках, что является общей чертой нейронной коммуникации.
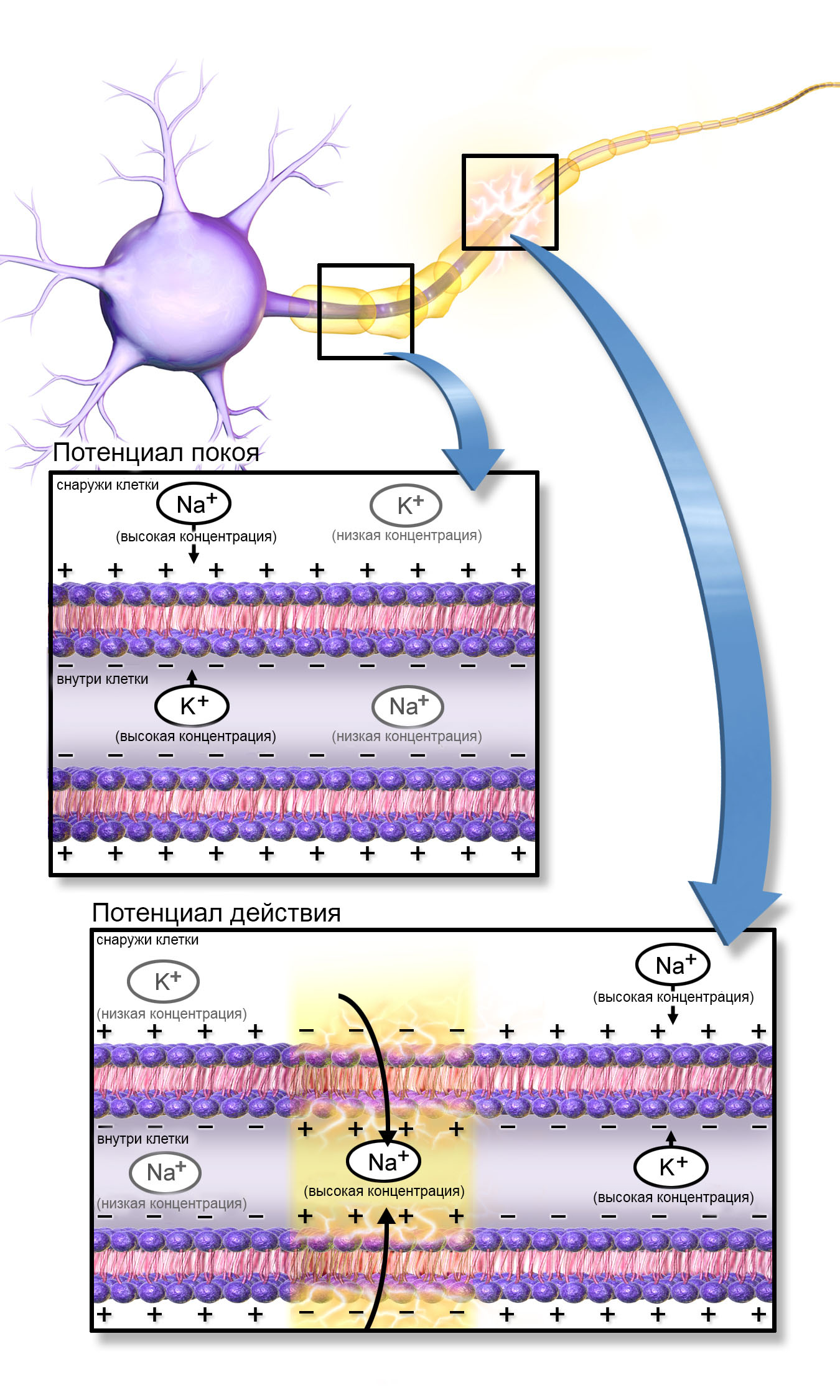
Потенциал покоя
Потенциал покоя (диффузионный потенциал) мембраны в норме существует в результате выхода из клетки положительно заряженных ионов (ионов калия (К+)), поддерживающих электрохимическое равновесие на уровне 75 мВ. В процессе деполяризации в клетку поступают положительно заряженные ионы (натрия (Na+)), что приводит к превышению нормального электрохимического потенциала покоя. Ионные каналы, расположенные в двойном липидном слое мембраны, открываются при помощи вольтаж-зависимого механизма и закрываются при помощи зависимого от времени механизма. Передача возбуждения к прилежащим участкам мембраны нейрона приводит к генерации потенциала действия при превышении порогового значения деполяризации.
Синаптическая передача
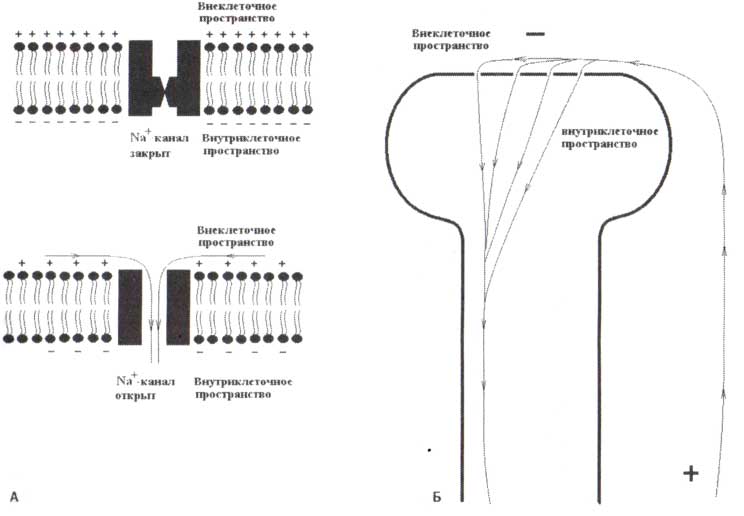
А. Схематическое представление закрытого и открытого дендритного Na-канала. Б. Стрелки показывают ионные токи во внеклеточном и внутриклеточном пространстве. Обратите внимание, что поток ионов (электрический ток) образует электрический диполь с положительным полюсом у основания дендрита.
Однако именно синаптический (возникающий на уровне синапсов) потенциал служит наиболее важным источником экстрацеллюлярных потоков, приводящих к генерации потенциала на ЭЭГ.
Синаптическая передача обеспечивает связь между нейронами, в связи с этим наблюдается взаимосвязанное изменение биопотенциалов в разных участках головного мозга.
Когда нейрон разряжается, он посылает потенциал действия через аксон к другой клетке, названной постсинаптическим нейроном. В синапсе, контакте между этими двумя нейронами, потенциал действия высвобождает медиатор. Медиатор распространяется в синаптическую щель и связывается с лиганд-зависимыми ионными каналами на постсинаптической мембране. Канал открывается и позволяет току катионов или анионов (в зависимости от типа нейронов — возбуждающего или тормозного) воздействовать на дендрит. Так как суммарный заряд дендрита равен нулю, ионный ток, направленный внутрь, на его постсинаптической мембране компенсируется ионным током, направленным наружу в противоположной части дендрита. Этот поток ионов внутрь дендрита и наружу производит маленький электрический ток (рис. 4).
- Возбуждающие постсинаптические потенциалы (ВПСП) распространяются по направлению внутрь клетки (из экстрацеллюлярного в интрацеллюлярное пространство) к другим компонентам клетки (sinks) посредством ионов натрия или кальция.
- Тормозные постсинаптические потенциалы (ТПСП) распространяются по направлению изнутри кнаружи (из интрацеллюлярного в экстрацеллюлярное пространство) в противоположном направлении (source), с участием ионов хлора или калия.
Эти суммированные потенциалы больше по продолжительности, чем потенциал действия, и определяют появление большинства колебаний, регистрируемых на ЭЭГ. Ствол мозга и таламус выполняют роль подкорковых генераторов, синхронизирующих популяции нейронов неокортекса как в норме (например, элементы сна), так и при патологии (например, генерализованные комплексы спайк—волна).
Нейроны коры большого мозга
Проводимость характеризует процесс перехода электрического тока от генератора в головном мозге на регистрирующий электрод.
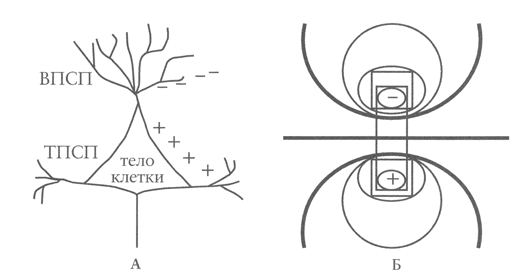
(А) Пирамидная клетка с возбуждающими постсинаптическими потенциалами (ВПСП) и тормозными постсинаптическими потенциалами (ТПСП). (Б) Диполь, описывающий разделение разнонаправленно заряженных полей
Нейроны коры большого мозга служат основным источником электрической активности на ЭЭГ. Пирамидные клетки вносят основной вклад в появление синаптического потенциала, формирующего картину ЭЭГ (рис. 5, А). Эти нейроны ориентированы в перпендикулярном направлении по отношению к поверхности коры и располагаются в слоях коры III, IV и VI. Генерируемая ими величина потенциала, достаточная для регистрации электрической активности на поверхности головы, требует вовлечения зон > 6 см2, хотя, вероятно, для регистрации большинства интериктальных эпилептиформных разрядов на поверхностной ЭЭГ (регистрируемой с поверхности головы) требуется участие нейрональных областей > 10 см2, так как кости черепа уменьшают проведение электрических сигналов. Все генераторы имеют позитивно и негативно направленные поля, выполняющие функцию диполя (рис. 5, Б). ЭЭГ отражает постоянное и изменяющееся напряжение электрических полей, варьирующее при различной локализации электродов на поверхности головы.
Электроэнцефалограмма
Скальповая (поверхностная) ЭЭГ регистрирует разницу электрических потенциалов между двумя участками на поверхности головы, расположенными над корой большого мозга (покрывающими соответствующие участки коры мозга), локализованными максимально близко к регистрирующему электроду. При рутинной ЭЭГ электрические потенциалы регистрируются непосредственно с поверхности головы, и регистрируемая активность на ЭЭГ проходит анализ частоты, амплитуды, морфологии и топографии колебаний. Однако большая часть коры больших полушарий человека скрыта глубоко под поверхностью скальпа и дополнительно отражает двумерные проекции трехмерного источника, что создает сложности при локализации генератора при анализе поверхностной ЭЭГ. Кроме того, электрическая активность, регистрируемая с поверхности головы, представляет собой суммарную синхронизированную активность больших популяций нейронов, которая создает корковый потенциал, и может не отображать небольшие интериктальные или иктальные источники патологической активности.