В данном обзоре рассмотрены различные теории систем, которые лежат в основе мыслительной деятельности и функционирования головного мозга.
Функциональная структура головного мозга может рассматриваться на разных уровнях в зависимости от масштаба. Каждый уровень формирует функциональные системы, которые обладают специфическими свойствами.
Микроуровень
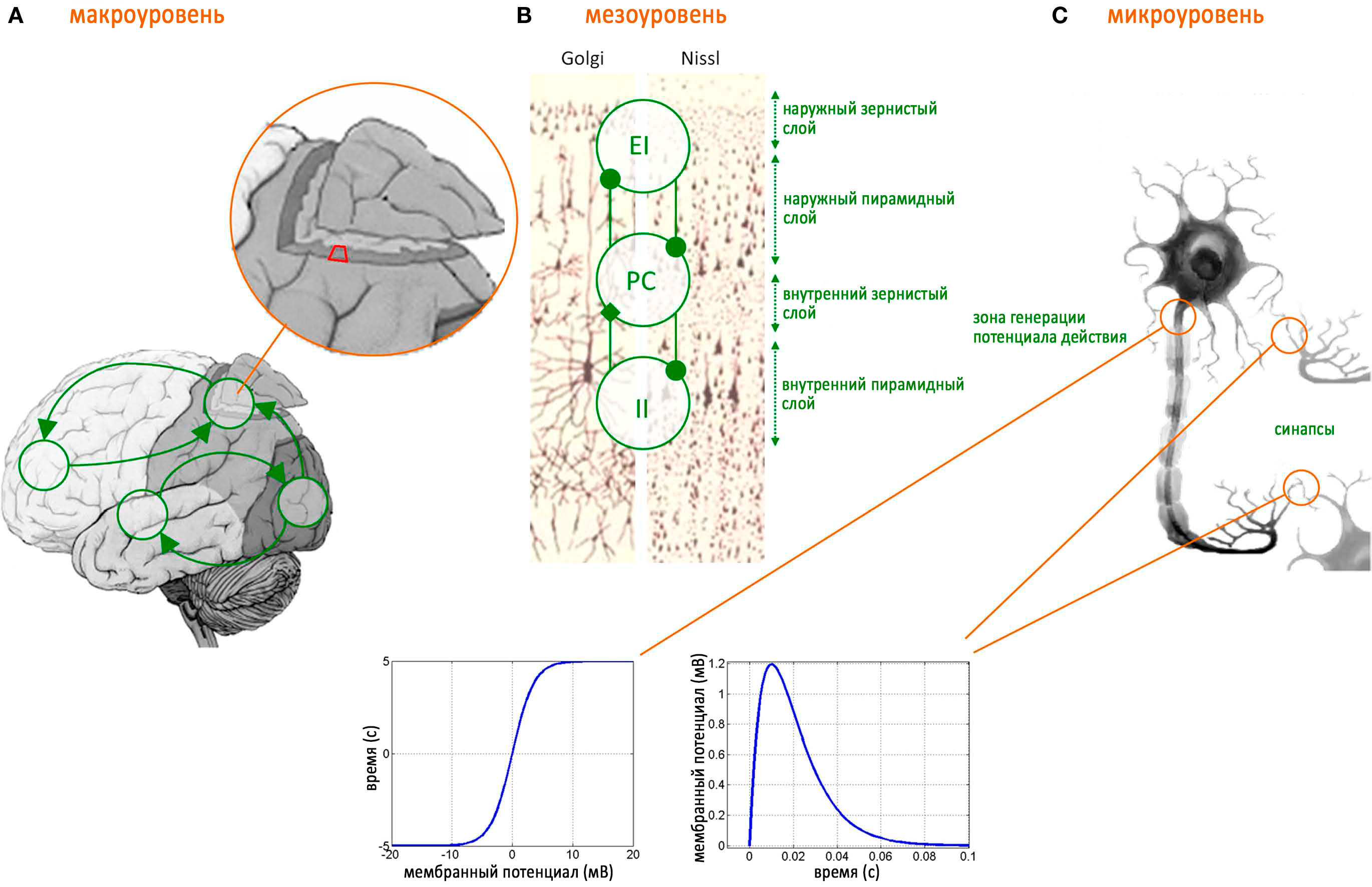
Уровень структурно-функциональной организации, на котором происходит обмен информацией между одиночными нейронами. Затрагивает молекулярные и синаптические механизмы генерации биоэлектрической активности.
Активность на микроуровне вызвана генерацией потенциала действия, в результате изменения заряда мембраны. Нейроны генерируют множественные потенциалы – пачки спайков (англ. spike trains), которые являются основой для нейронного кодирования и передачи информации в мозге. Они формируют все виды паттернов и на более высоких уровнях.
Одиночные нейроны способны генерировать подпороговые колебания мембранного потенциала (англ. subthreshold membrane potential oscillations), что не сопровождается формированием потенциала действия. Это происходит, когда постсинаптический потенциал не смог достигнуть порога возбудимости нейрона и не активировал его, либо это может быть связано со спонтанными изменениям потенциала нейронов.
По возбудимости нейроны можно подразделить на I (нейроны-интеграторы) и II (нейроны-резонаторы) классы.
У нейронов I класса малое смещение мембранного равновесия затухает экспоненциально (монотонно), они могут генерировать потенциалы действия с произвольно с низкой частотой в зависимости от силы входных импульсов, тогда как у нейронов II класса, возмущение состояния равновесия мембранного потенциала вызывает ответ в виде затухающих колебаний, так как при повторяющейся подпороговой стимуляции они весьма чувствительны к частоте следования импульсов. Для них, в отличие от интеграторов, увеличение частоты стимуляции может затормозить, или даже прекратить генерацию ответного потенциала действия. Нейроны II класса также более склонны к проявлению подпороговых колебаний мембранного потенциала.
Мезоуровень
Мезоуровень структурно-функциональной организации головного является промежуточным и обеспечивает связь между отдельными нейронами и всем мозгом. Образован кортикальными нейронными сетями, размером от нескольких миллиметров до сантиметров. Временное разрешение для регистрации активности на этому уровне составляет от миллисекунд до секунд. На данном уровне происходит взаимодействие больших нейронных ансамблей между собой. Активность на этом уровне организации можно измерить такими методами, как LFP (потенциалы локального поля), ЭКоГ (электрокортикография), ЭЭГ (электроэнцефалография) или МЭГ (магнитоэнцефалография). Часто данное понятие «мезо» можно встретить как просто указание на то, что область интересов находится между «микро» и «макро».
Макроуровень
Высший уровень, или макроуровень обеспечивает реализацию основных функций головного мозга. Например, обработка сенсорной информации, принятие решений, моторная и двигательная активность. Временные задержки играют здесь важную роль. Поскольку все области мозга связаны двунаправленно, эти связи между областями мозга образуют петли обратной связи. Петли положительной обратной связи имеют тенденцию вызывать ритмическую активность, когда частота обратно пропорциональна времени задержки. Примером являются связи между таламусом и корой – таламо-кортикальные связи. Эта таламо-кортикальная сеть способна генерировать колебательную активность, известную как таламо-кортикальный резонанс. Таламо-кортикальная система мозга играет важную роль в генерации альфа-активности. В модели сети всего мозга с реалистичной анатомической связностью и задержками распространения между областями мозга, колебания в диапазоне бета-частот возникают в результате частичной синхронизации подмножеств областей мозга, колеблющихся в гамма-диапазоне (генерируемых в мезоскопическом уровне).
Ритмическая активность в основе функциональной организации мозга
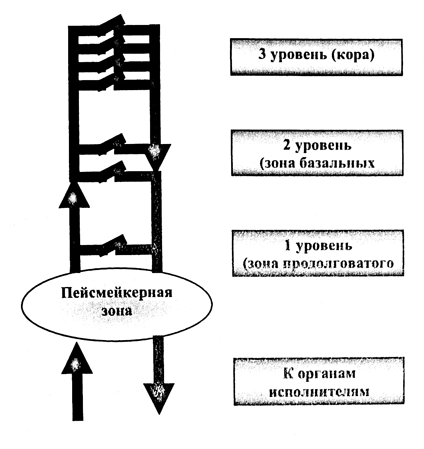
Головной мозг представляет собой сложноорганизованную структуру, состоящую их огромного числа нервных клеток, и синаптических связей. В единой нервной сети, эти структуры могут рассматриваться как система переключателей, поддерживающих, перенаправляющих или прерывающих основное нервное возбуждение. Однако, для своего функционирования, эта система нуждается в первичном импульсе, который в центральной нервной системе вырабатывается в специальном образовании, т.н. пейсмейкерной зоне.
Согласно современным данным, она расположена на уровне ростральных отделов ствола, в зоне олив и ретикулярной формации, но не тождественна им. В этой зоне находятся группы нервных клеток, способных к ритмичному спонтанному возбуждению. Над ними расположены ядерные образования продолговатого мозга (зона переключения нейронов I-го уровня), таламус и подкорковые образования (зона переключения нейронов II-го уровня), лимбическая система, интимно связанная таламусом и подкорковыми образованиями и непосредственно кора больших полушарий.
Пейсмейкерные клетки обеспечивают постоянную ритмическую стимуляцию всех структур головного мозга (а также поддержание исходного мышечного тонуса), посылая импульсы с частотой 6-8 Гц. Схематично, данную организацию можно представить как систему разноуровневых переключателей (рис. 1), количество которых возрастает от уровня продолговатого мозга к коре. Каждый переключатель, при своей активации вносит свою «помеху» в общий электрический сигнал пейсмейкерного пула. Большее количество переключателей соответственно больше деформируют основной сигнал.
Таким образом, вышележащие отделы, трансформируют основной ритм по принципу, чем больше включенных нейронных структур, тем выше частота и ниже амплитуда колебаний.
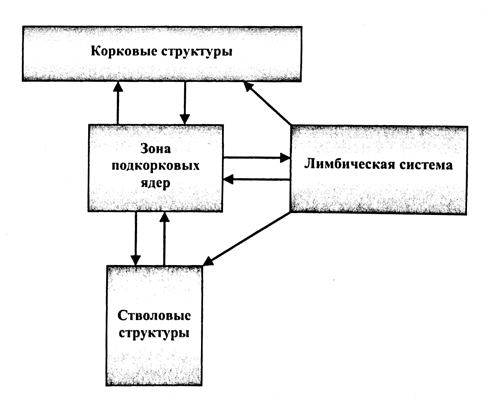
Изучение особенностей записи ЭЭГ с корковых и подкорковых образований, позволило канадскому ученому В. Пенфилду разработать теоретическую концепцию функциональной организации головного мозга на основе т.н. центрэнцефалической системы. В ее основе лежит положение о том, что активация мозговых структур производится из единого центра активации, расположенного в ростральных отделах продолговатого мозга. Активированные вышестоящие образования в свою очередь оказывают воздействие на центр активации по механизму обратной связи (рис. 2).
Более поздние исследования подтвердили эту теорию, доказав слабую биоэлектрическую активность в корковых структурах, функционально разобщенных с подкорковыми образованиями. Помимо этого, была установлена роль подкорковых образований, преимущественно таламуса, в регуляции и интеграции биоэлектрической активности корковых структур.
Теория функциональных систем П. К. Анохина
В данном случае функциональная система это объединение центральных и периферических элементов организма, которое позволяет ему (организму) решать ту или иную задачу, выполнять ту или иную функцию.
«Принцип функциональной системы» — объединение частных механизмов организма в целостную систему приспособительного поведенческого акта, создание «интегративной единицы».
Выделяются два типа функциональных систем:1
- Системы первого типа обеспечивают гомеостаз за счёт внутренних (уже имеющихся) ресурсов организма, не выходя за его пределы (напр. кровяное давление);
- Системы второго типа поддерживают гомеостаз за счёт изменения поведения, взаимодействия с внешним миром, и лежат в основе различных типов поведения.
Данная теория рассматривает поведенческий акт – т.е. совокупность процессов, протекающих от восприятия раздражителя до оценки последствий – следующим образом:
Афферентный синтез
- Любое возбуждение в центральной нервной системе существует во взаимодействии с другими возбуждениями: головной мозг проводит анализ этих возбуждений. Синтез детерминируют следующие факторы:
- Мотивация
- Пусковая афферентация (возбуждения, вызываемые условными и безусловными раздражителями)
- Обстановочная афферентация (возбуждение от привычности обстановки, вызывающей рефлекс, и динамические стереотипы)
- Память (видовая и индивидуальная)
Принятие решения
- Формирование акцептора результата действия (создание идеального образа цели и его удержание; предположительно, на физиологическом уровне представляет собой циркулирующее в кольце интернейронов возбуждение)
Эфферентный синтез (или же стадия программы действия; интеграция соматических и вегетативных возбуждений в единый поведенческий акт. Действие сформировано, но не проявляется внешне)
- Действие (выполнение программы поведения)
- Оценка результата действия
На этом этапе идёт сравнение реально выполняемого действия с идеальным образом, созданным на этапе формирования акцептора результата действия (происходит обратная афферентация); на основании результатов сравнения действие или корректируется, или прекращается.
- Удовлетворение потребности (санкционирующая прекращение деятельности стадия)
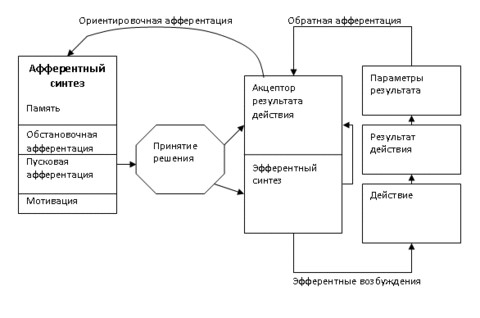
Таким образом, поведение каждого организма рассматривается как совокупность множества узкоспециализированных функциональных систем (как врожденных, так и приобретенных), обеспечивающих приспособление к условиям окружающего мира.234
На биологическом уровне, каждая функциональная система представляет собой коллектив нервных клеток, управляемый из головного мозга при участии периферических элементов. При этом совокупность всех функциональных систем, взаимодействующих между собой, представляет собой человеческое “Я”, его самосознание, как таковое.
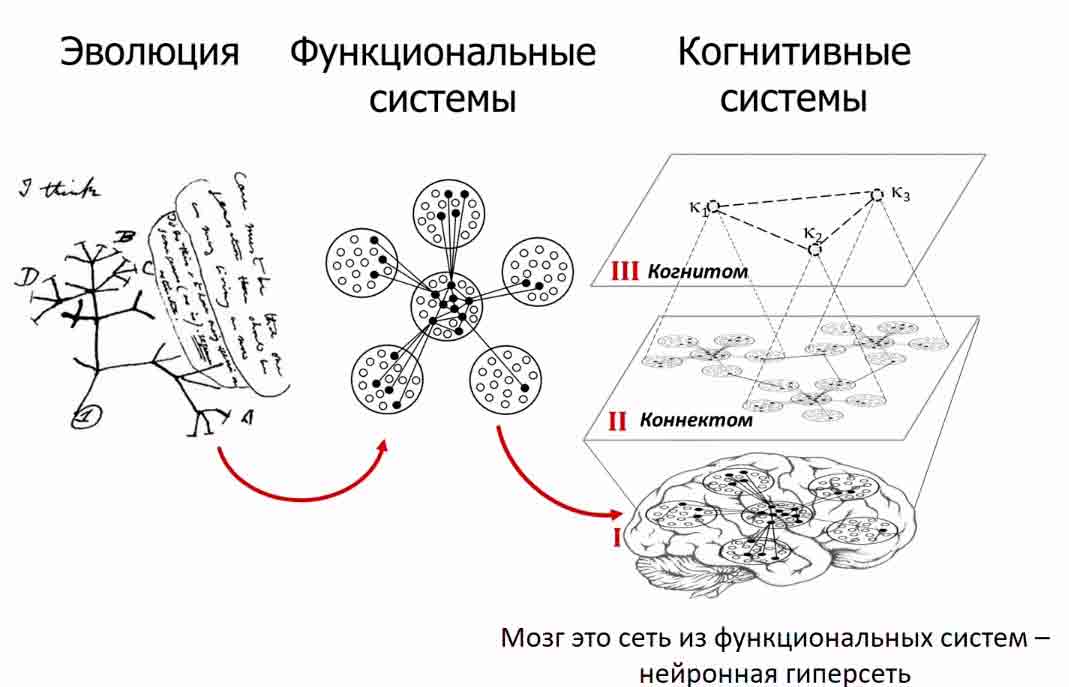
Элементы функциональных систем распределены по всему головному мозгу и отделены, но не обособлены друг от друга. Они находятся в тесном взаимодействии, и активация одной способна вызвать активацию ряда других. Сеть таких взаимосвязанных нейронов (которые являются базовой единицей функциональной системы) называется кооперативной группой, когнитивной группой или когом и содержит в себе знания и опыт индивида, как врожденные, так и приобретенные. Вся совокупность когов называется когнитόм.
Дальнейшее развитие теории функциональных систем привело к созданию теории нейронных гиперсетей. Согласно данной теории, любой разум формализуем, т.е. может быть описан как нейронная гиперсеть – сеть, в которой вершинами выступают сети из функционально связанных нейронов. Главным положением данной теории является возможность описания разума, как органической и математической структуры, а также исследование его с использованием методов экспериментальной и теоретической физики, подходов теории графов и статистической механики.
Основными положениями и утверждениями данной теории являются:
- Разум реален и является нейронной гиперсетью, т.е. высокопорядковой структурой мозга, которая опосредует каузальные (причинно-следственные) соотношения организма со средой;
- Для описания элементов и свойств этой системы вводятся новые фундаментальные понятия:
- КоГ (CoG) – от англ. cognitive group, рус. когнитивная группа – элементы разума, формирующие его «зернистую» структуру. Они опосредуют информационные соотношения целостного когнитивного агента с его средой. Т.о. разум гранулярен и состоит из простейших ментальных элементов;
- ЛиГ (link of CoGs) – от англ. link of cognitive group, рус. сеть когнитивных групп – это устойчивые когнитивные связи между отдельными когами. Они отражают причинные связи элементов и процессов в среде и в соотношениях когнитивного агента с ней. Т.о. отдельные элементы разума связаны друг с другом;
- Когнитом (cognitome) – от англ. сognitive network, рус. когнитивная сеть – единая когнитивная сеть из когов и лигов, которая является носителем всего субъективного опыта. Т.о. разум системен и образует уникальную структуру личности.
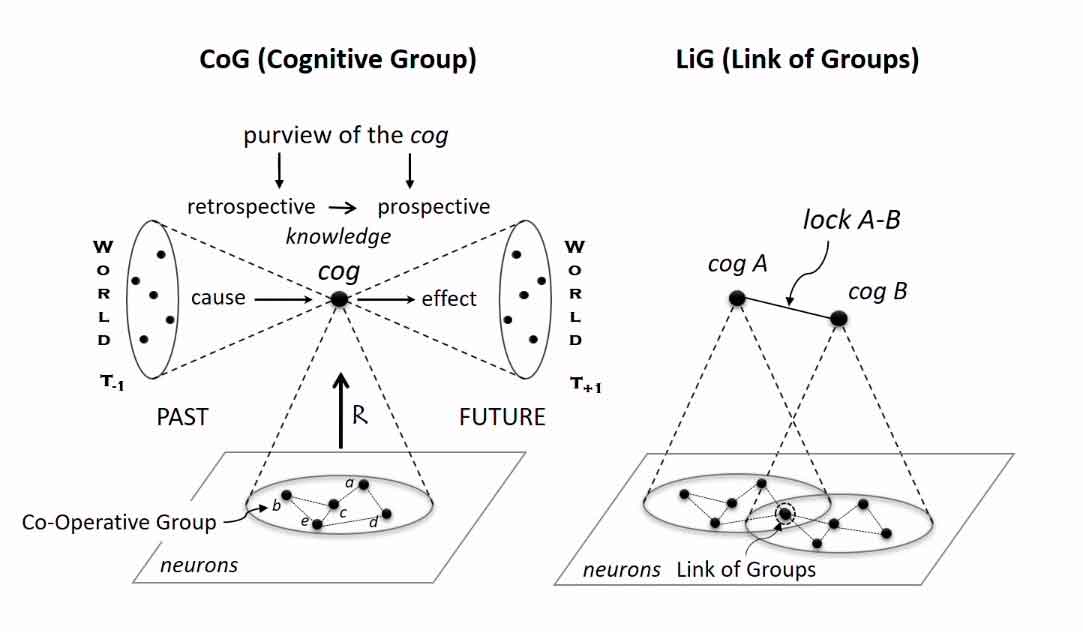
В левой части рисунка изображена реакция организма на раздражитель из внешнего мира. Информация о совокупности неких событий в окружающей действительности, совпадает с имеющимся в головном мозге когом. Соответственно, при поступлении в нервную ткань сигнала активируется система связей между клетками, вызывая в сознании субъекта определенную реакцию. Необходимо понимать, что, во-первых, каждой группе клеток соответствует одно конкретное событие, а во-вторых эта взаимосвязь индивидуальна для каждого субъекта и формируется на основании его личного опыта. Кроме того, данный факт позволяет делать вывод от обратного – активация в головном мозге определенного кога свидетельствует о возникновении в мире определенного события.
Однако, несмотря на связь «одно событие – один ког», связи «один ког – одна реакция» не существует. В связи с морфологическими особенностями строения нейронов, при активации одной группы (одного кога) происходит иррадиация возбуждения на соседние клетки, что приводит к активации других когов и проявлению набора поведенческих реакций. Тем не менее, данный процесс сужает количество возможных ответных действий организма до обозримого числа.
Кроме того, данный процесс формирует ожидания организма относительно будущих событий, т.е. формирует способность к прогнозированию за счет сравнения поступающих сигналов с имеющимися когами.
В правой части иллюстрации изображен принцип работы лигов. Его суть заключается в том, что при активации кога, повышается вероятность проявления связанной с ним группы за счет передачи возбуждения по лигу. Это приводит к формированию механизма ассоциаций.
Т.о. теория нейронных гиперсетей рассматривает мозг, как многоуровневую сеть взаимосвязанных вычислительных структур, распределенных по всему объему головного мозга.
Существенным вопросом в современной нейронауке является то, почему нейроны – и только они – являются одновременно и обычной нервной клеткой, и носителем субъективной информации.
Функциональные блоки мозга А. Р. Лурии
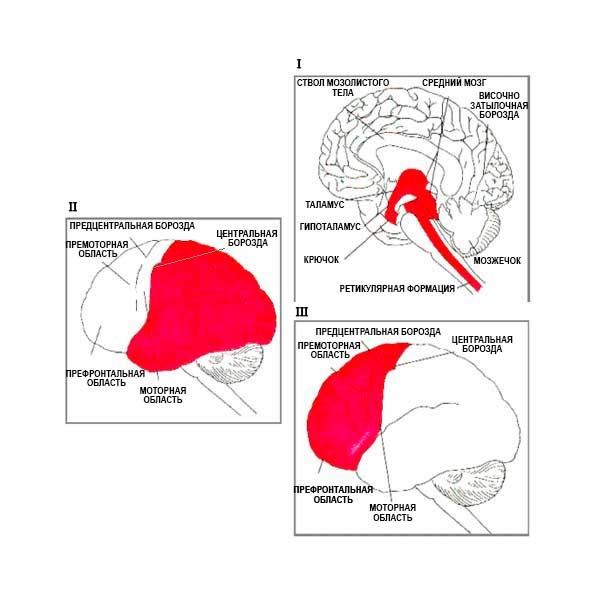
Лурия утверждал, что психические процессы человека представляют собой сложные функциональные системы, в которые вовлечены группы областей мозга, работающих согласованно, каждая из которых вносит уникальный вклад в организацию функциональной системы.
Таким образом, Лурия обозначил три основные функциональные единицы мозга, необходимые для психических процессов человека в целом и сознательной деятельности в частности:
- блок для регулировки тонуса или бодрствования;
- блок для получения, обработки и хранения информации;
- блок для программирования, регулирования и проверки умственной деятельности.
Каждая из этих трех единиц имеет иерархическую структуру, состоящую из трех корковых зон, основанных одна на другой: первичная (проекционная) область, которая получает импульсы от периферии или посылающая импульсы на периферию, вторичная (проекционно-ассоциативная) область, куда поступает, обрабатывается информация и разрабатываются программы, а также третичные (зоны перекрытия) области, которые отвечают за самые сложные формы умственной деятельности, требующие согласованного вовлечения многих областей коры головного мозга.
Двухкомпонентная модель биоэлектрогенеза
➥ Основная статья: Синхронизирующая и десинхронизирующая системы мозга
В суммарной ЭЭГ отражается активность системы таламокортикальных нейронов. Но эта система не существует изолированно, а находится под постоянным влиянием других систем мозга5. В результате суммарная ЭЭГ отражает общий уровень функциональной активности всего головного мозга. На этом основании рассмотрение механизмов биоэлектрогенеза теоретически следует строить, исходя из оценки влияния всех структур мозга на функциональное состояние коры. Практически это невозможно. Первичной моделью описания механизмов биоэлектрогенеза является так называемая двухкомпонентная модель, которая рассматривает ЭЭГ как результат интерференции активности двух систем мозга: синхронизирующей и десинхронизирующей. В синхронизирующую систему включают структуры, снижающие уровень бодрствования, поэтому она называется также сомногенной системой. Сихронизирующая система включает главным образом неспецифические таламические ядра, структуры нижних отделов моста и некоторые ядра продолговатого мозга. На ЭЭГ снижение уровня бодрствования проявляется генерацией медленноволновой активности. Чем ниже уровень бодрствования, тем ниже частота суммарной активности и, как правило, больше амплитуда. «Замедление» доминирующей активности в паттерне суммарной ЭЭГ в нейрофизиологии обозначается как синхронизация.
Стимуляция структур восходящей активирующей системы вызывает реакцию пробуждения (arousal). При этом на ЭЭГ регистрируется переход к генерации активности высокой частоты. Такая перестройка обозначается как десинхронизация биоэлектрической активности.
Функции тормозных и возбуждающих систем мозга
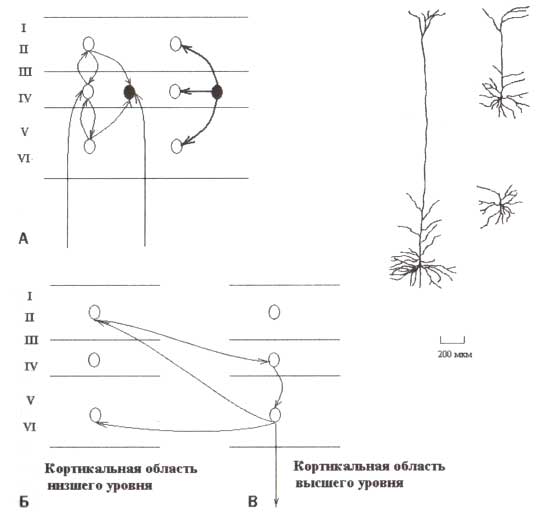
Электроды, расположенные на скальпе, регистрируют потенциалы главным образом от корковых областей, расположенных около поверхности головы. Ниже будет детально описана анатомия и физиология корковых генераторов, которые производят потенциалы, распространяющиеся к поверхности головы и измеряющиеся скальповыми электродами в форме электроэнцефалограммы. Нейроны с их аксонами, сомами и дендритами — первичные кандидаты на роль таких генераторов потенциалов. Один из основных принципов корковой организации — существование в коре возбуждающих и тормозных нейронов (рис. 7). Эти типы нейронов предназначены природой для формирования сложных действий в корковых нейронных сетях:
- Обнаружение совпадения необходимо для того, чтобы увеличить отношение сигнал/шум, обрабатывающегося нейронной сетью. Когда входы к ассоциативной коре от первичных и вторичных сенсорных областей, извлекающих различные особенности стимула. совпадают во времени, датчик совпадения сигнализирует, что эти входы не случайны, а принадлежат тому же самому стимулу.
- Пространственная фильтрация выполняет ту же функцию (увеличение отношения «сигнал/шум»), что и обнаружение совпадения, но имеет другой механизм. Этот механизм основывается на пространственной (а не временной, как в случае обнаружения совпадения) интеграции раздражений электрической активности мозга, прибывающих в определенный нейрон от пространственно распределенной сети. Возбуждающие и тормозные входы формируют сложный пространственный фильтр. Например, рецепторные поля сложных клеток в первичной зрительной коре можно было бы рассматривать как пространственные фильтры Габора, в то время как соответствующее действие в зрительной системе рассматривают как пространственную фильтрацию.
- Реверберация сигнала в нейронных сетях необходима, чтобы сохранять сигнал в краткосрочной памяти. Теоретическая концепция реверберации была известна с 1940-х, но только недавно было получено прямое нейрофизиологическое доказательство. Критическая структурная предпосылка для реверберации — существование рекуррентных цепей в нейронных сетях мозга.
- Сравнение двух входов — произведенного ожидаемой моделью поведения и произведенного текущей ситуацией — играет критическую роль в сенсорных и исполнительных системах. В сенсорной системе это необходимо для обнаружения изменений, то есть для обнаружения любого изменения в монотонном сенсорном возбуждении. В исполнительной системе действие сравнения необходимо для обнаружения несоответствия между запланированным действием и действием, которое было фактически выполнено субъектом. Обнаружение этого изменения, в свою очередь, служит для того, чтобы контролировать поведение в общем, и для тою, чтобы исправлять ошибки в частности.
В общих чертах процессы возбуждения и торможения в коре сбалансированы. Как возбуждающие, так и тормозные клетки коры получают возбуждающие входы от таламуса. Таламус, в свою очередь, рассматривается как корковые ворота, так как все сенсорные пути, берущие начало в сенсорных органах (например, сетчатка, кохлеарные ядра), проходят через релейные ядра таламуса. Нерелейные ядра таламуса получают информацию от базальных ганглиев, так же как и си первичных сенсорных корковых областей, и посылают проекции к остальной части коры. Наибольшее количество входов от таламуса в кору распределено в пределах срединных корковых областей. Кроме того, возбуждающие и тормозные нейроны получают прямые проекции от корковых областей, расположенных на более низком уровне корковой иерархии, и обратные проекции от корковых областей, расположенных на более высоком уровне корковой иерархии.
Возбуждающие нейроны в пределах локального коркового модуля связаны друге другом, формируя цепь реверберации, которая оказывается ответственной за поддержание локальной электрической активности на какое-то время дольше, чем длительность действия стимула. Этот процесс функционально основывается на следующих процессах локальных прямой и обратной передачи: когда группа возбужденных нейронов посылает возбуждающие потенциалы соседним нейронам, они разряжаются с некоторой задержкой и посылают возбуждающие потенциалы назад к первоначально задействованной группе клеток, активизируя их снова (рекуррентно). Эта последовательность прямой и обратной передачи возбуждения ведет к лавинообразному возбуждению и ответственна за поддержание активации в пределах локальной цепи.
Тормозные нейроны получают возбуждающие импульсы непосредственно от таламуса и через окружающие возбуждающие нейроны. Тормозные нейроны имеют обратные проекции на локальные возбуждающие нейроны. Тормозные нейроны выполняют по крайней мере три важных функции. Во-первых, текущее торможение не позволяет «перевозбуждаться» локальным корковым цепям. Во-вторых, пространственный паттерн распределения тормозных клеток формирует сложные воспринимающие поля нейронов в коре. В-третьих, комбинация торможения и возбуждения может формировать различные типы операций сравнения в пределах коры. Возбуждающие нейроны в коре разделены на два типа: пирамидные клетки и звездчатые клетки. Примеры этих двух типов возбуждающих нейронов представлены на рис. 7. Короткая длина дендритов позволяет звездчатым нейронам обрабатывать поступающие сигналы быстро и с высокой временной точностью. Пирамидный нейрон отличается от звездчатого нейрона наличием очень длинного, так называемого апикального дендрита, который в 6 раз длиннее, чем базальные дендриты. Апикальный дендрит ориентирован перпендикулярно к корковой поверхности.
Этот тип ориентации дендритов называют вертикальной организацией в отличие от горизонтальной (вдоль корковой поверхности). Короткие дендриты пирамидных клеток простираются от тела нейрона практически во всех направлениях и называются базальными дендритами. Возможно, апикальные дендриты играют особую роль в обработке информации в пределах корковых нейронных сетей. Эта функция все еще требует объяснения, но мы знаем наверняка, что длинные апикальные дендриты пирамидных нейронов коры играют критическую роль в генерации ЭЭГ, регистрируемой со скальпа.
Разнообразие функций коркового торможения достигается разнообразными классами тормозных интернейронов: корзинчатыми, клеткими-канделябрами, нейронами двойного букета, нейроглиаформными клетками. Аксоны этих интернейронов имеют высокоспецифичную геометрию и паттерны иннервации. Клеточные механизмы, которые определяют тормозные связи электрической активности головного мозга, только начинают становиться быть понятными.
Оценка особенностей системно-функциональной организации мозговой деятельности по ЭЭГ
Оценка уровня организации мыслительной деятельности по ЭЭГ подразумевает электроэнцефалографическую оценку организующей и контролирующей роли префронтальной коры. При этом может быть выделен электроэнцефалографический синдром гипофронтальности и гиперфронтальности.
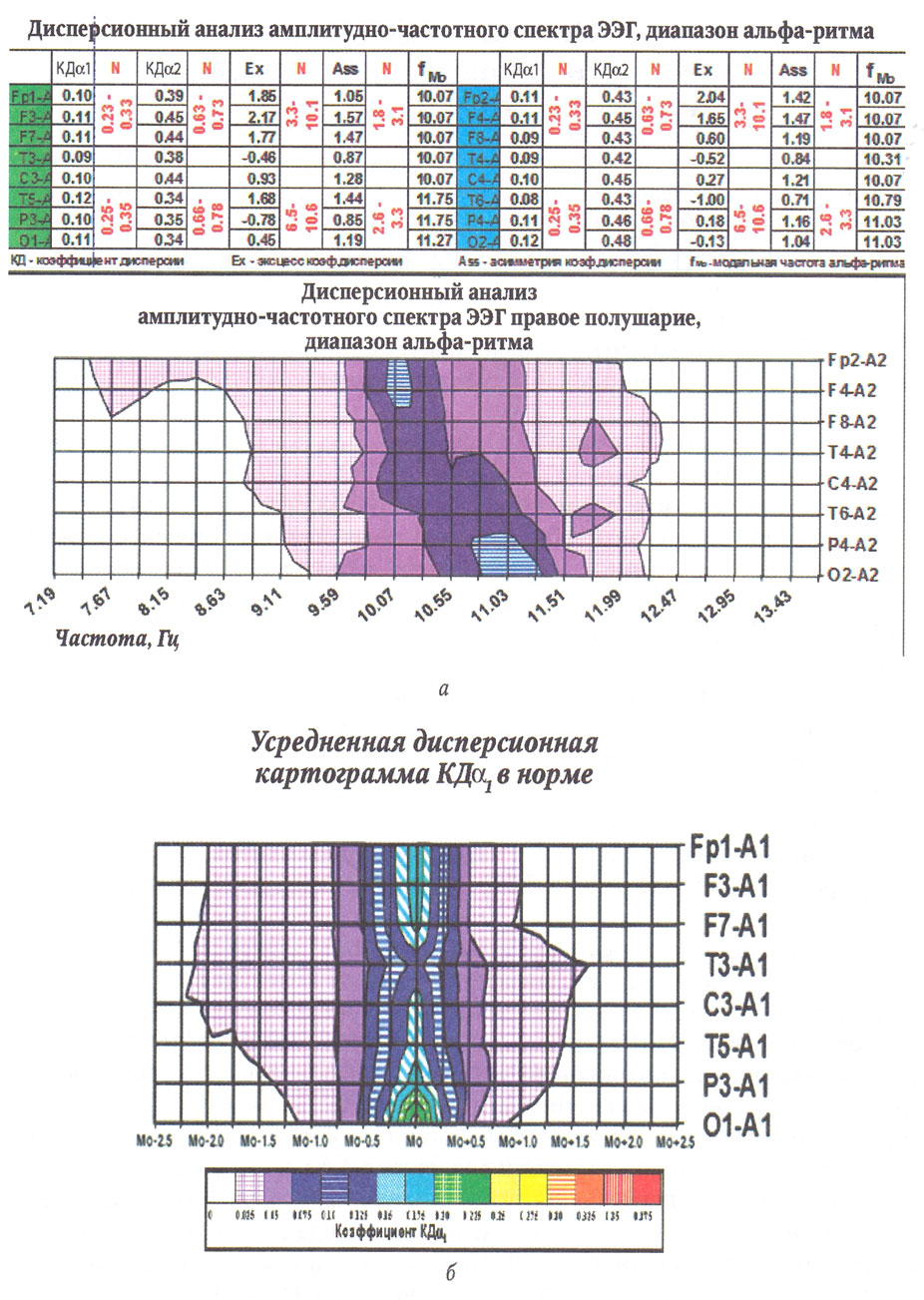
Для гипофронтальности характерными признаками являются:
- расширенная частотная представленность альфа-ритма на спектрах мощности, на дисперсионных картах, низкие значения коэффициента дисперсии по Росману67;
- пространственная девиация альфа-ритма со смещением в сторону замедления модальных значений в передних отделах по спектру мощности, показателям дисперсии или периодометрического анализа (рис.);
- снижение уровня межлобных связей по значениям когерентности при сопутствующих повышенных уровнях связей в центральных и теменных отделах;
- инверсия переднезаднего соотношения внутриполушарных связей за счет снижениея уровня в передних отделах или повышения его в задних;
- крайне низкий уровень лобно-затылочной синхронизации (по данным когерентного анализа).
Гиперфронтальность на ЭЭГ сопровождается:
- сужением частотной представленности альфа-ритма в виде мономодальности с очень узким диапазоном на спектрах мощности и с максимальной консолидацией в передних отделах на дисперсионных картах;
- пространственной моночастотностью суженного диапазона альфа-ритма;
- избыточно высоким уровнем синхронизации альфа-ритма по внутри-и межполушарным связям, включая лобно-затылочные пары.
Интерпретация состояния системно-функциональной организации мозговой деятельности по ЭЭГ подразумевает электроэнцефалографическую оценку организующей и контролирующей роли префронтальной коры. В основу оценочного критерия была положена гипотетическая модель функциональной организации вероятного информационного обмена между зонами мозга, где носителями кванта информации являются альфа-колебания.
Основанием для применения такой модели явился факт совпадения модальных значений с умеренной степенью консолидации частоты альфа-ритма в теменно-затылочной и лобной областях, установленной у 165 здоровых людей8, и частотной девиацией в сторону замедления у пациентов с СДВГ и парасомниями, у которых обнаружены признаки диссоциации уровня бодрствования9. Анализ большого количества электроэнцефалограмм у пациентов с аномалиями в психическом статусе, где был использован в качестве основного метода оценки пространственной девиации альфа-ритма дисперсионный анализ в сопоставлении с оценкой уровня внутримозговых связей по данным когерентного анализа ЭЭГ, показал две основные тенденции изменений. В одном варианте отмечалось расширение дисперсионной зоны альфа-ритма со смещением модальных значений в сторону замедления в передних отделах, с ослаблением частотной консолидации по данным дисперсионного анализа и снижение уровня связей в передних отделах мозга и по длинным ассоциативным путям, во втором отмечалась обратная картина, наблюдались избыточная лобно-затылочная консолидация альфа-ритма с выраженным сужением частотной зоны в передних отделах, а также избыточный уровень функциональных связей по данным когерентного анализа. Эти варианты изменений сформулированы как электроэнцефалографические синдромы гипофронтальности и гиперфронтальности.
По этим критериям ЭЭГ гипофронтальность выявляется у большинства пациентов с СДВГ, церебрастенией, посттравматической энцефалопатией, при парасомниях, соматоформных расстройствах и др.
Гиперфронтальность выявляется практически во всех наблюдениях с выраженными паническими атаками. Существенно реже она наблюдалась при СДВГ при парасомниях, соматоформных расстройствах, астеноневротиче-ских реакциях и только при условии наличия в клинике тревожного аффекта.
В ряде наблюдений бывает сложно в чистом в виде отнести изменения на ЭЭГ либо к гипофронтальности, либо к гиперфронтальности, так как, с одной стороны, имело место дисперсионное «размытие» частотного состава альфа-диапазона, с другой — наблюдается избыточная консолидация модальных значений одной из составляющих доминирующего альфа-ритма, или реже показатели когерентного анализа вступают в противоречие с картиной дисперсии альфа-ритма. Такая картина более характерна для смешанного варианта невротического паттерна ЭЭГ.
Читайте также
- Функциональная организация мозга
- Функциональная активность мозга
- Структурно-функциональная модель мозга
- Обратная задача ЭЭГ
Footnotes
- The theory of functional systems: General postulates and principles of dynamic organization. Konstantin V. Sudakov. Integrative Physiological and Behavioral Science volume 32, pages 392–414(1997) DOI: 10.1007/BF02688634
- Anokhin PK (1937): The functional system as the basis for the integration of nervous processes of embryogenesis. In: Proceedings of the VIth All-Union Congress of Physiologists, Biochemists and Pharmacologists (pp. 148–156). Tbilisi, Orgkomitet.
- Anokhin PK (1948): Systemogenesis as a general principle of evolutionary development. Bull. Experim. Biology and Medicine 26: 81–99.
- Anokhin PK (1968): Biology and Neurophysiology of the Conditioned Reflex. Moscow, Meditsina.
- Электроэнцефалография : руководство / М. В. Александров, Л. Б. Иванов, С. А. Лытаев [и др.] / под ред. М. В. Александрова. — 3-е изд., перераб. и доп. — Санкт-Петербург: СпецЛит, 2020. — 224 с.
- Росман С. В. Диагностические возможности дисперсионного картирования электроэнцефалограммы // Психическое здоровье. — 2013. – №6. -С. 64-69.
- Росман С. В. Применение дисперсионного картирования альфа-ритма в раннем выявлении психических болезней //Врач. — 2013. — № 8. — С. 79— 82.
- Росман С. В. Диагностические возможности дисперсионного картирования электроэнцефалограммы // Психическое здоровье. — 2013. – №6. -С. 64-69.
- Иванов Л. Б., Стрекалина Н. Н., Чулкова Н. Ю., Будкевиг А. В. Варианты пространственного распределения альфа-активности в зависимости от формы аффективных расстройств // Функциональная диагностика. — 2009. — № 1. – С. 41-50.